

Activin A induces Langerhans cell differentiation in vitro and in human skin explants
- PMID: 18813341
- PMCID: PMC2533393
- DOI: 10.1371/journal.pone.0003271
Activin A induces Langerhans cell differentiation in vitro and in human skin explants
Abstract
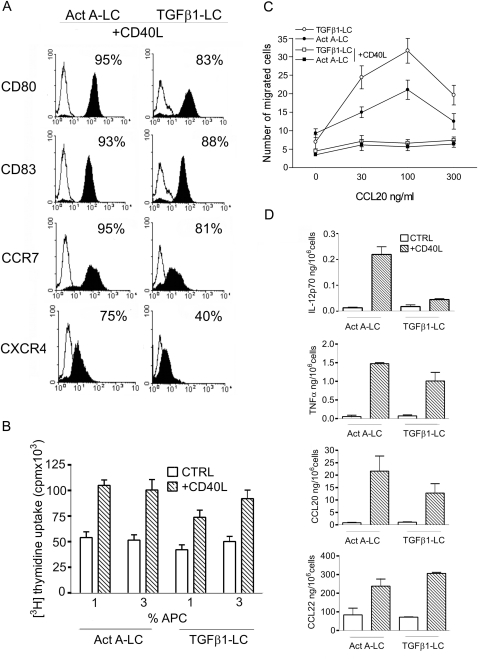
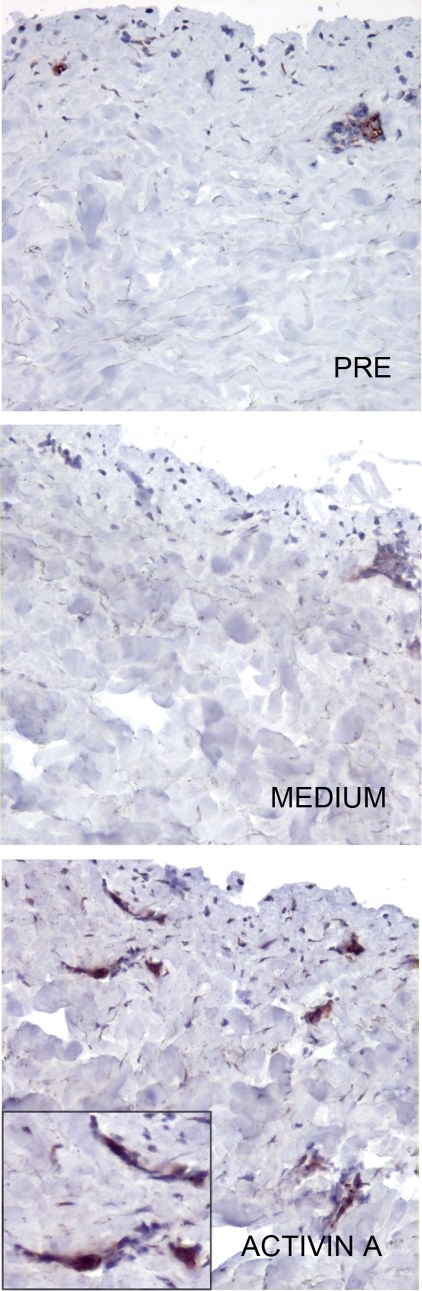
Langerhans cells (LC) represent a well characterized subset of dendritic cells located in the epidermis of skin and mucosae. In vivo, they originate from resident and blood-borne precursors in the presence of keratinocyte-derived TGFbeta. In vitro, LC can be generated from monocytes in the presence of GM-CSF, IL-4 and TGFbeta. However, the signals that induce LC during an inflammatory reaction are not fully investigated. Here we report that Activin A, a TGFbeta family member induced by pro-inflammatory cytokines and involved in skin morphogenesis and wound healing, induces the differentiation of human monocytes into LC in the absence of TGFbeta. Activin A-induced LC are Langerin+, Birbeck granules+, E-cadherin+, CLA+ and CCR6+ and possess typical APC functions. In human skin explants, intradermal injection of Activin A increased the number of CD1a+ and Langerin+ cells in both the epidermis and dermis by promoting the differentiation of resident precursor cells. High levels of Activin A were present in the upper epidermal layers and in the dermis of Lichen Planus biopsies in association with a marked infiltration of CD1a+ and Langerin+ cells. This study reports that Activin A induces the differentiation of circulating CD14+ cells into LC. Since Activin A is abundantly produced during inflammatory conditions which are also characterized by increased numbers of LC, we propose that this cytokine represents a new pathway, alternative to TGFbeta, responsible for LC differentiation during inflammatory/autoimmune conditions.
Conflict of interest statement
Figures



References
-
- Banchereau J, Briere F, Caux C, Davoust J, Lebecque S, et al. Immunobiology of dendritic cells. Annu Rev Immunol. 2000;18:767–811. - PubMed
-
- Bell D, Young JW, Banchereau J. Dendritic cells. Adv Immunol. 1999;72:255–324. - PubMed
-
- Steinman RM, Banchereau J. Taking dendritic cells into medicine. Nature. 2007;449:419–426. - PubMed
-
- Stingl G, Katz SI, Shevach EM, Rosenthal AS, Green I. Analogous functions of macrophages and Langerhans cells in the initiation in the immune response. J Invest Dermatol. 1978;71:59–64. - PubMed
-
- Strobl H, Riedl E, Bello-Fernandez C, Knapp W. Epidermal Langerhans cell development and differentiation. Immunobiology. 1998;198:588–605. - PubMed
