

Hsf1 is required for the nuclear translocation of p53 tumor suppressor
- PMID: 18813348
- PMCID: PMC2546585
- DOI: 10.1593/neo.08430
Hsf1 is required for the nuclear translocation of p53 tumor suppressor
Abstract
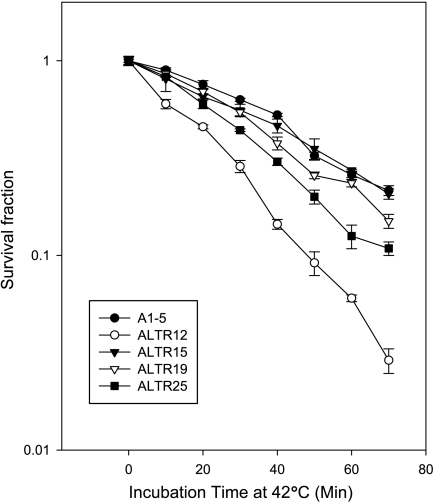
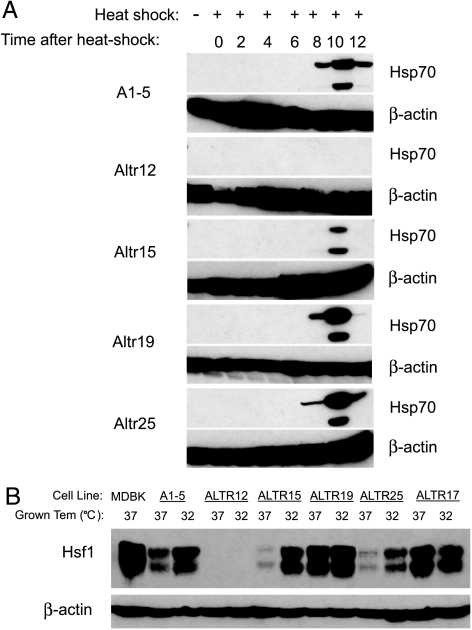
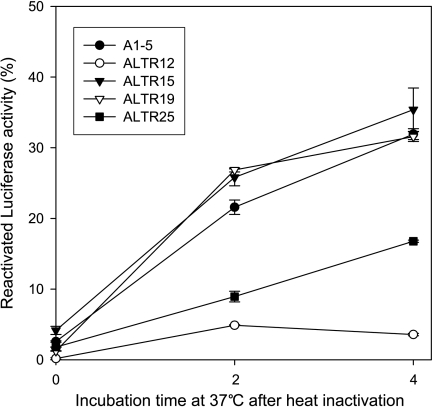
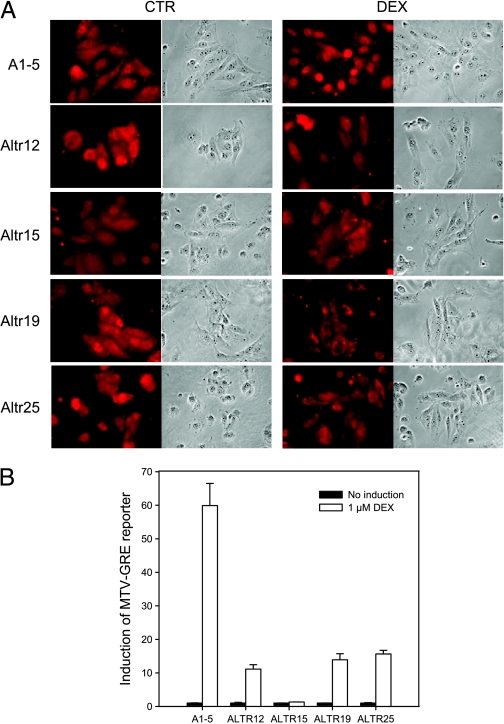
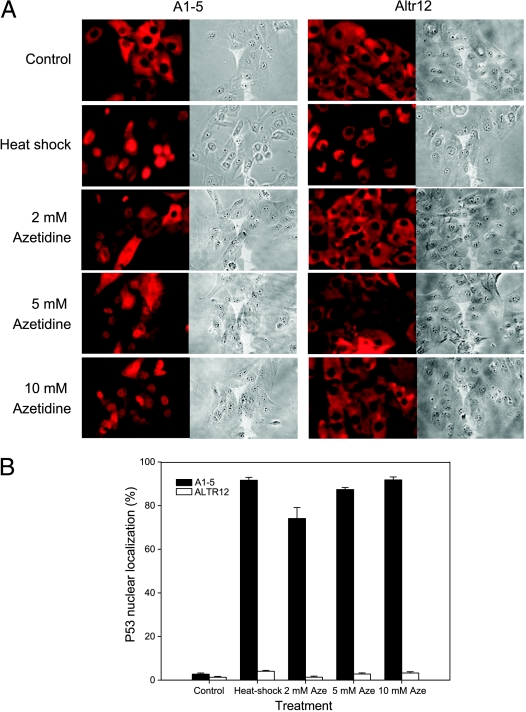
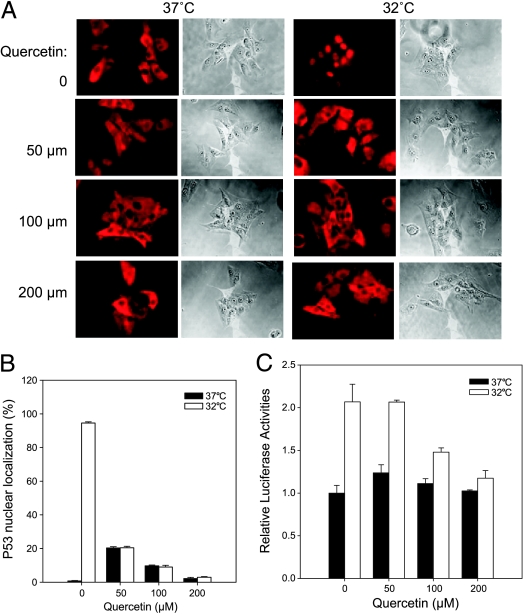
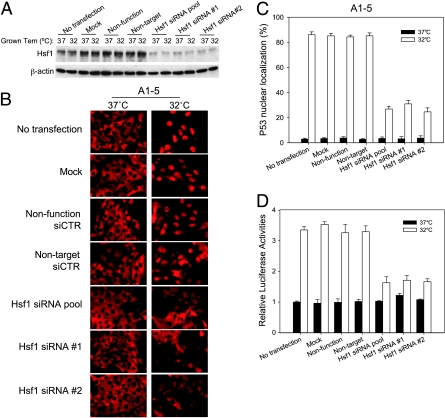
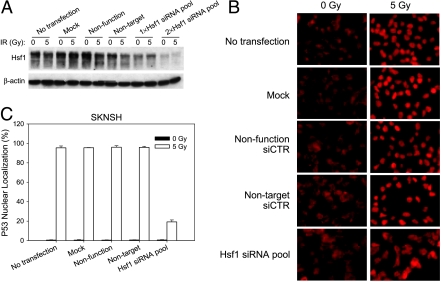
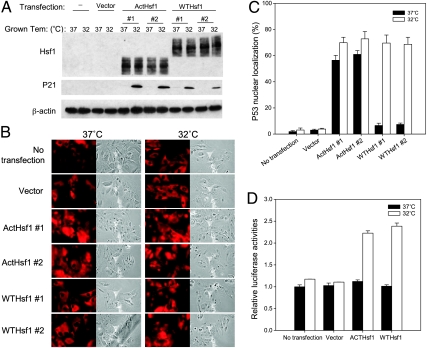
Although the p53 tumor suppressor is most frequently inactivated by genetic mutations, exclusion from the nucleus is also seen in human tumors. We have begun to examine p53 nuclear importation by isolating a series of mutant cells in which the temperature-sensitive murine p53(Val135) mutant is sequestered in the cytoplasm. We previously showed that that three of them (ALTR12, ALTR19, and ALTR25) constituted a single complementation group. Here, we found that ALTR12 cells are more sensitive to heat stress than either ALTR19 or ALTR25 and that there was a complete lack of induction of Hsp70 in response to heat shock. Western blot analysis showed no expression of the Hsf1 transcription factor, and neither heat shock nor azetidine could induce p53 nuclear localization in ALTR12 cells but did in parental A1-5 cells. Suppression of Hsf1 in A1-5 cells with quercetin or an Hsf1 siRNA reduced p53 nuclear importation and inhibited p53-mediated activation of a p21 reporter. Most convincingly, p53 nuclear importation could be restored in ALTR12 cells by introducing an exogenous Hsf1 gene. Collectively, our result suggests that Hsf1 is required for p53 nuclear importation and activation and implies that heat shock factors play a role in the regulation of p53.
Figures
References
-
- Komarova EA, Zelnick CR, Chin D, Zeremski M, Gleiberman AS, Bacus SS, Gudkov AV. Intracellular localization of p53 tumor suppressor protein in gamma-irradiated cells is cell cycle regulated and determined by the nucleus. Cancer Res. 1997;57:5217–5220. - PubMed
-
- Fritsche M, Haessler C, Brandner G. Induction of nuclear accumulation of the tumor-suppressor protein p53 by DNA-damaging agents. Oncogene. 1993;8:307–318. [Erratum appears in Oncogene 1993 8 (9), 2605] - PubMed
-
- Liang SH, Clarke MF. A bipartite nuclear localization signal is required for p53 nuclear import regulated by a carboxyl-terminal domain. J Biol Chem. 1999;274:32699–32703. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous