

doi: 10.1039/b803512a.
Epub 2008 Jun 27.
Fe(II)/alpha-ketoglutarate hydroxylases involved in nucleobase, nucleoside, nucleotide, and chromatin metabolism
Affiliations
- PMID: 18813363
- PMCID: PMC2907160
- DOI: 10.1039/b803512a
Item in Clipboard
Fe(II)/alpha-ketoglutarate hydroxylases involved in nucleobase, nucleoside, nucleotide, and chromatin metabolism
Dalton Trans.
.
Abstract
Fe(II)/alpha-ketoglutarate-dependent hydroxylases uniformly possess a double-stranded beta-helix fold with two conserved histidines and one carboxylate coordinating their mononuclear ferrous ions. Oxidative decomposition of the alpha-keto acid is proposed to generate a ferryl-oxo intermediate capable of hydroxylating unactivated carbon atoms in a myriad of substrates. This Perspective focuses on a subgroup of these enzymes that are involved in pyrimidine salvage, purine decomposition, nucleoside and nucleotide hydroxylation, DNA/RNA repair, and chromatin modification. The varied reaction schemes are presented, and selected structural and kinetic information is summarized.
Figures
General reaction of FeII/αKG hydroxylases
Pyrimidine salvage pathway. Selected fungi are capable of salvaging thymine from dTMP by a series of reactions catalyzed by (1) pyrimidine deoxynucleoside-2′-hydroxylase, (2) nucleoside hydrolase or phosphorylase, (3) T7H, (4) uracil-5-carboxylate decarboxylase (also named iso-orotate decarboxylase), and (5) uracil phosphoribosyltransferase. The resulting UMP is available for conversion to other nucleotides by standard reactions.
Structure of XanA according to a homology model. (A) The predicted overall fold of the XanA protein is depicted as a green ribbon. (B) A blow-up of the proposed active site. In both panels, FeII is an orange sphere, αKG has yellow carbon atoms, metal ligands have blue carbon atoms, a cofactor-stabilizing residue (Arg 352) is shown with orange carbon atoms, and other postulated active site residues are indicated with white carbon atoms.
Structure of XanA according to a homology model. (A) The predicted overall fold of the XanA protein is depicted as a green ribbon. (B) A blow-up of the proposed active site. In both panels, FeII is an orange sphere, αKG has yellow carbon atoms, metal ligands have blue carbon atoms, a cofactor-stabilizing residue (Arg 352) is shown with orange carbon atoms, and other postulated active site residues are indicated with white carbon atoms.
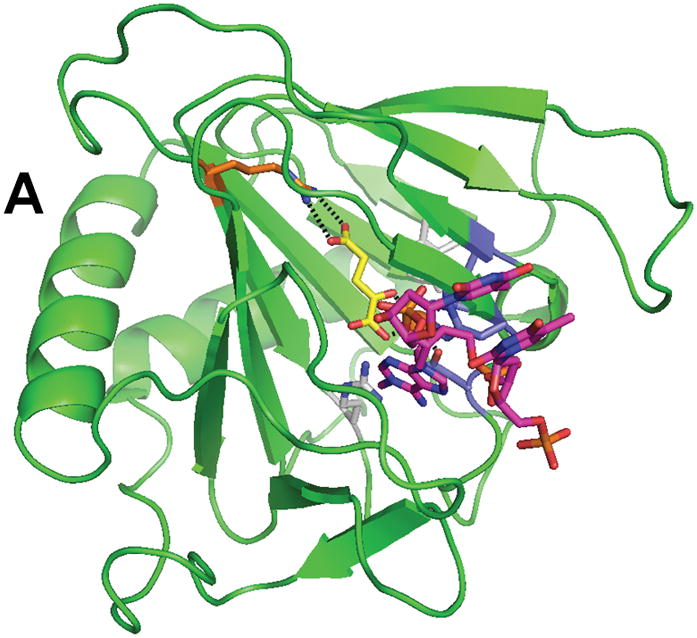
Structure of AlkB (PDB 2fd8). (A) The overall fold of AlkB. (B) The active site residues of AlkB. The coloring scheme is the same as for Figure 3, and the bound T-1meA-T substrate is shown with pink carbon atoms (the substrate is abbreviated to just 1meA in B).
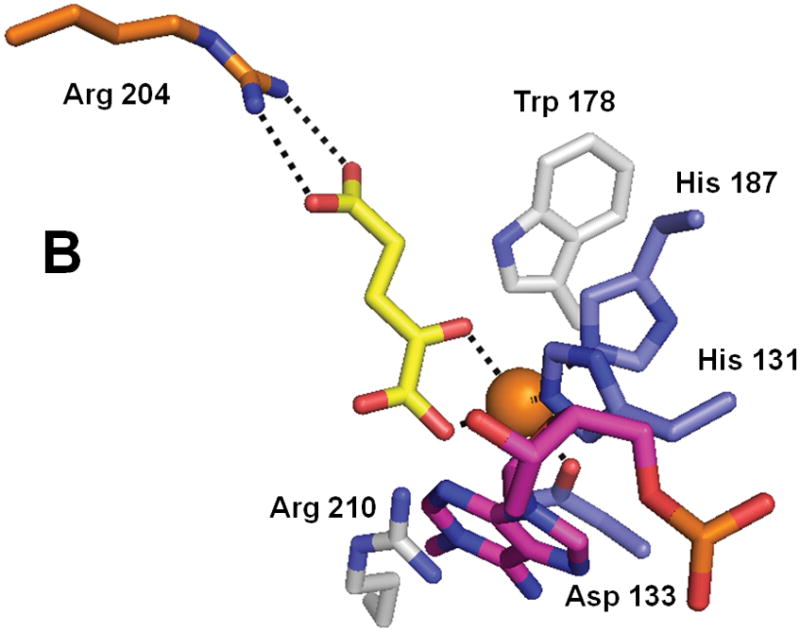
Structure of AlkB (PDB 2fd8). (A) The overall fold of AlkB. (B) The active site residues of AlkB. The coloring scheme is the same as for Figure 3, and the bound T-1meA-T substrate is shown with pink carbon atoms (the substrate is abbreviated to just 1meA in B).

Structure of ABH2 (PDB 3buc). (A) The overall fold of ABH2 with tethered DNA containing 1meA. (B) The active site residues of ABH2 with 1meA. To reduce oxygen reactivity, the active site FeII was replaced by MnII (depicted as a gray sphere). The rest of the coloring scheme is the same as above.

Structure of ABH2 (PDB 3buc). (A) The overall fold of ABH2 with tethered DNA containing 1meA. (B) The active site residues of ABH2 with 1meA. To reduce oxygen reactivity, the active site FeII was replaced by MnII (depicted as a gray sphere). The rest of the coloring scheme is the same as above.
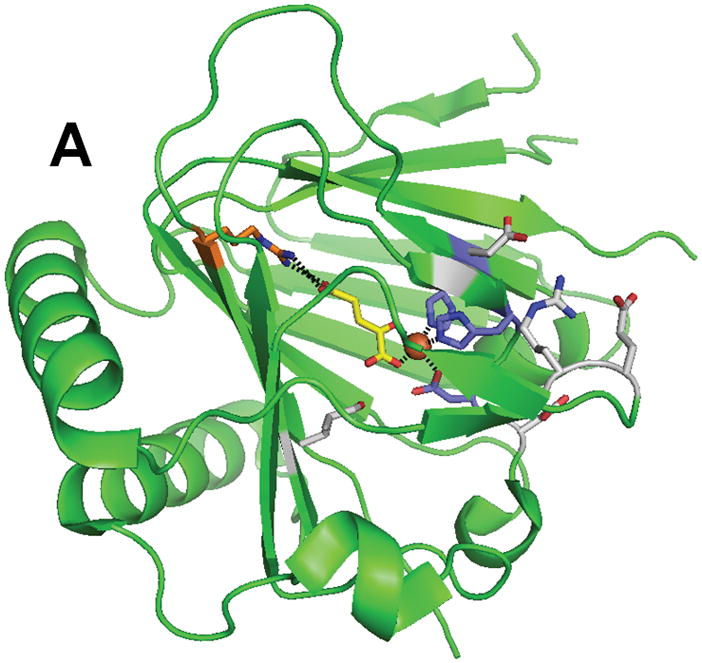
Structure of ABH3 (PDB 2iuw). (A) The overall fold of ABH3. (B) A blow-up of the ABH3 active site with coloring as indicated above. Note that Leu177 an oxidized species.
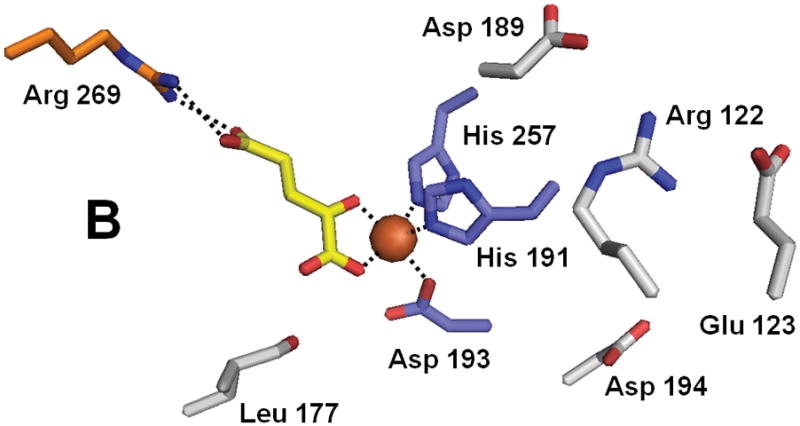
Structure of ABH3 (PDB 2iuw). (A) The overall fold of ABH3. (B) A blow-up of the ABH3 active site with coloring as indicated above. Note that Leu177 an oxidized species.
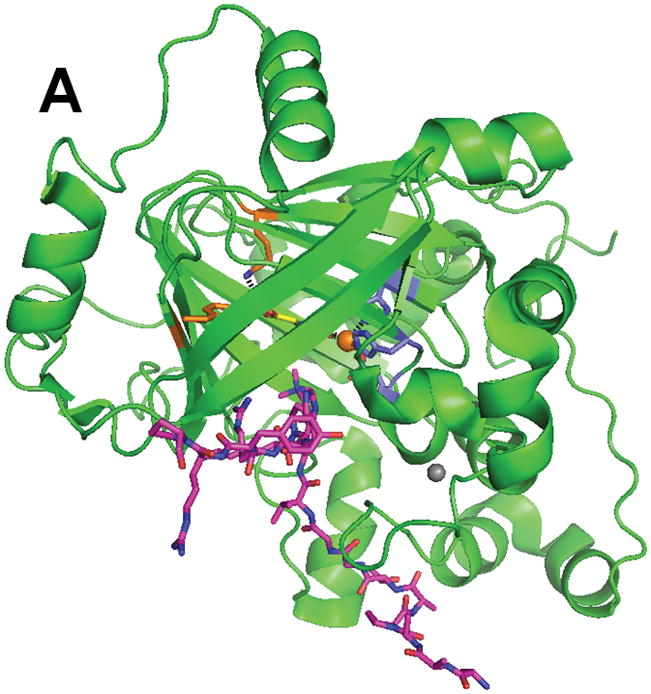
Structure of the histone demethylase JMJD2A (PDB 2p5b). (A) The overall view of the histone demethylase, with bound zinc shown as a grey sphere, the peptide substrate shown with pink carbons, and the αKG homologue N-oxalylglycine replacing the cosubstrate. (B) The active site of JMJD2A with just the trimethyl-Lys portion of the substrate shown. The coloring scheme is the same above.
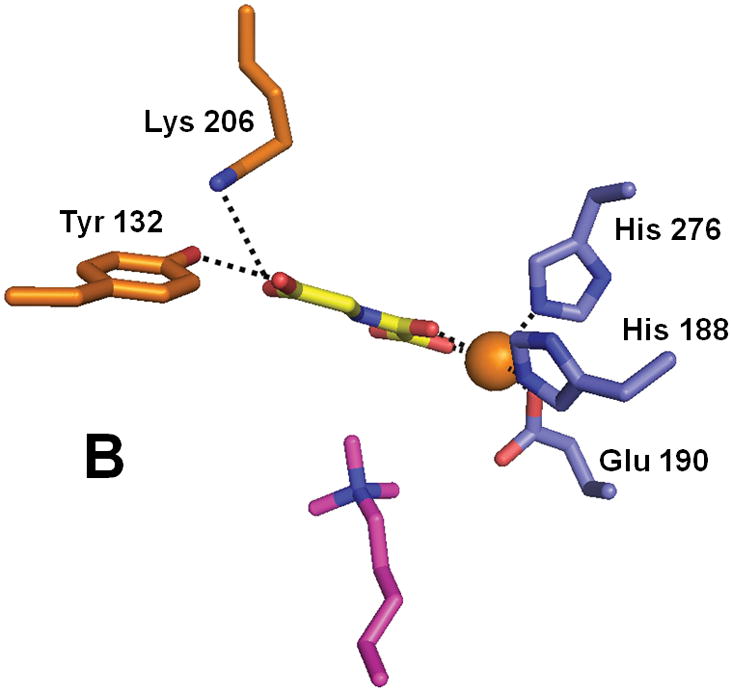
Structure of the histone demethylase JMJD2A (PDB 2p5b). (A) The overall view of the histone demethylase, with bound zinc shown as a grey sphere, the peptide substrate shown with pink carbons, and the αKG homologue N-oxalylglycine replacing the cosubstrate. (B) The active site of JMJD2A with just the trimethyl-Lys portion of the substrate shown. The coloring scheme is the same above.
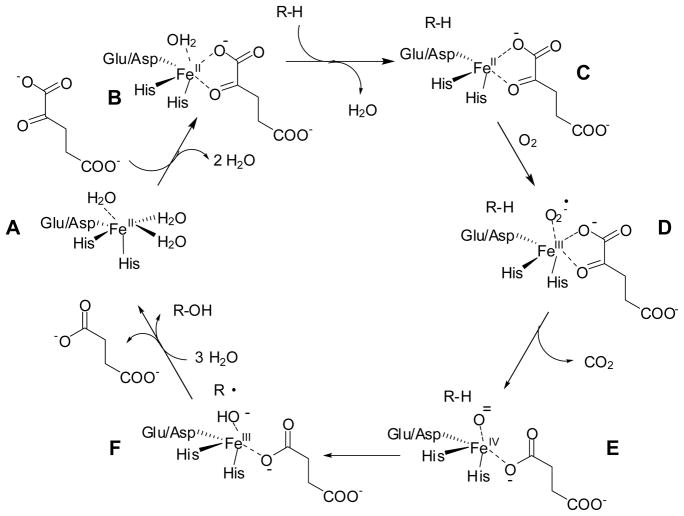
General mechanism of FeII/αKG hydroxylases
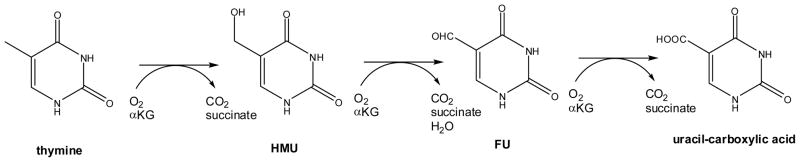
Reactions of thymine 7-hydroxylase
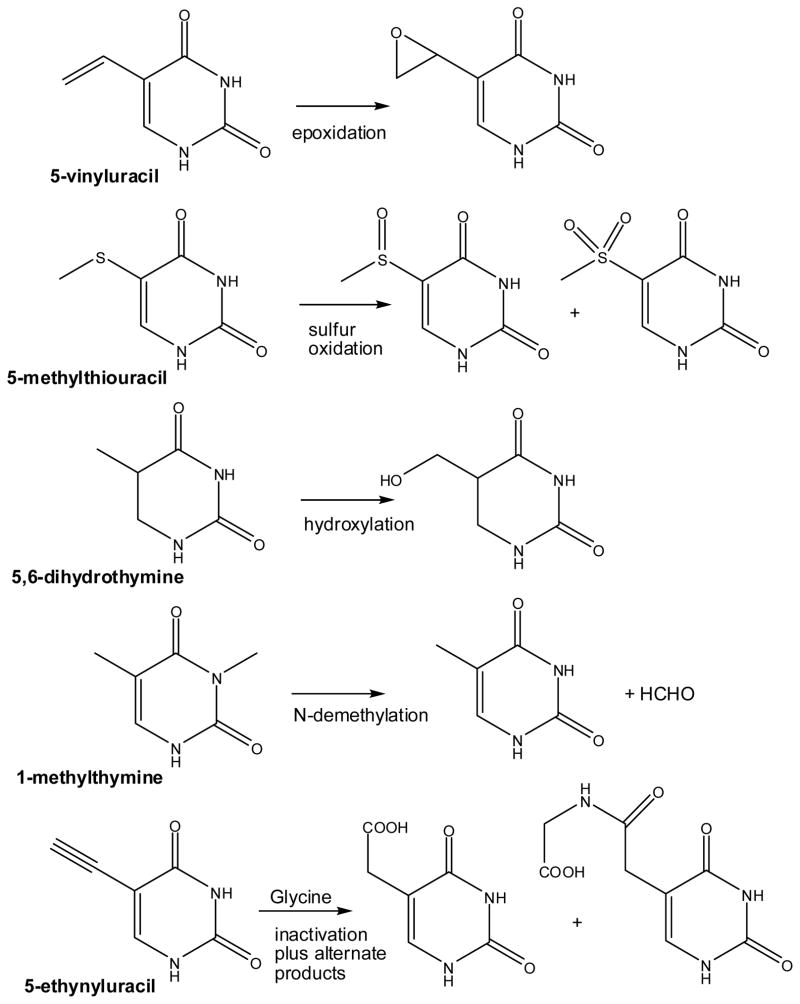
Diversity of alternative reactions catalyzed by thymine 7-hydroxylase
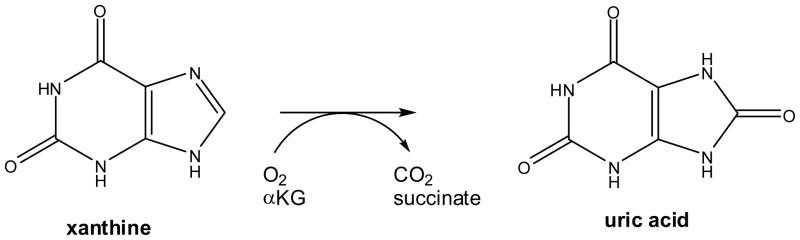
Reaction of FeII/αKG-dependent xanthine hydroxylase

Reaction of pyrimidine deoxyribonucleoside 2′-hydroxylase with thymidine (X = CH3) or deoxyuridine (X = H)

Reaction of deoxyuridine (uridine) 1′-hydroxylase
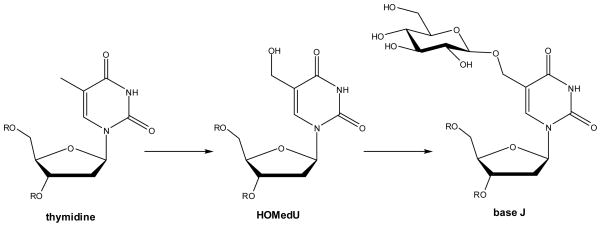
Biosynthesis of base J in kinetoplastids

Reactions of AlkB with 1meA and 3meC in DNA
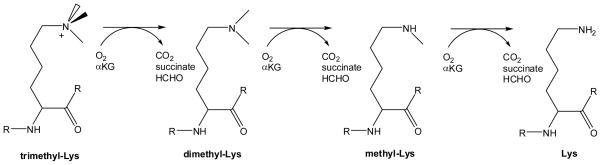
Reactions of methyl-Lys demethylases involved in removal of methyl groups from specific Lys residues of certain histones

Reactions of methyl-Arg demethylases involved in removal of methyl groups from specific Arg residues in selected histones
References
-
- Hausinger RP. Crit Rev Biochem Mol Biol. 2004;39:21–68. - PubMed
-
- Purpero V, Moran GR. J Biol Inorg Chem. 2007;12:587–601. - PubMed
-
- Loenarz C, Schofield CJ. Nature Chemical Biology. 2008;4:152–156. - PubMed
-
- Clifton IJ, McDonough MA, Ehrismann D, Kershaw NJ, Granatino N, Schofield CJ. J Inorg Biochem. 2006;100:644–669. - PubMed
-
- Hanauske-Abel HM, Günzler V. J Theor Biol. 1982;94:421–455. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources