

Posttranslational regulation of NF-YA modulates NF-Y transcriptional activity
- PMID: 18815279
- PMCID: PMC2592644
- DOI: 10.1091/mbc.e08-03-0295
Posttranslational regulation of NF-YA modulates NF-Y transcriptional activity
Abstract
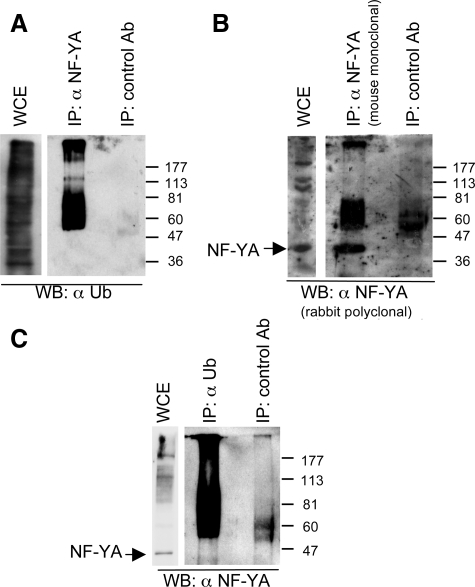
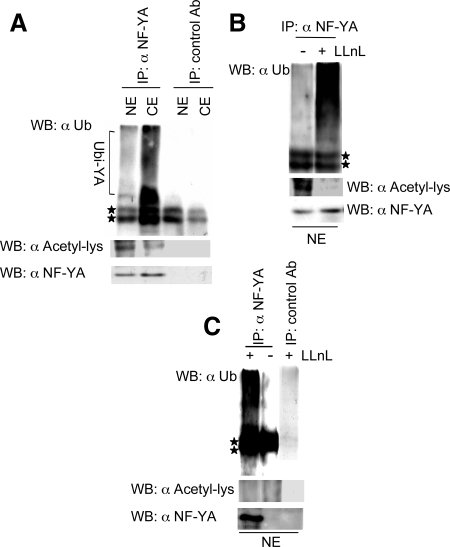
NF-Y binds to CCAAT motifs in the promoter region of a variety of genes involved in cell cycle progression. The NF-Y complex comprises three subunits, NF-YA, -YB, and -YC, all required for DNA binding. Expression of NF-YA fluctuates during the cell cycle and is down-regulated in postmitotic cells, indicating its role as the regulatory subunit of the complex. Control of NF-YA accumulation is posttranscriptional, NF-YA mRNA being relatively constant. Here we show that the levels of NF-YA protein are regulated posttranslationally by ubiquitylation and acetylation. A NF-YA protein carrying four mutated lysines in the C-terminal domain is more stable than the wild-type form, indicating that these lysines are ubiquitylated Two of the lysines are acetylated in vitro by p300, suggesting a competition between ubiquitylation and acetylation of overlapping residues. Interestingly, overexpression of a degradation-resistant NF-YA protein leads to sustained expression of mitotic cyclin complexes and increased cell proliferation, indicating that a tight regulation of NF-YA levels contributes to regulate NF-Y activity.
Figures









References
-
- Bhattacharya A., Deng J. M., Zhang Z., Behringer R., de Crombrugghe B., Maity S. N. The B subunit of the CCAAT box binding transcription factor complex CBF/NF-Y) is essential for early mouse development and cell proliferation. Cancer Res. 2003;63:8167–8172. - PubMed
-
- Bolognese F., Wasner F., Dohna C. L., Gurtner A., Ronchi A., Muller H., Manni I., Mossner J., Piaggio G., Mantovani R., Engeland K. The cyclin B2 promoter depends on NF-Y, a trimer whose CCAAT-binding activity is cell cycle regulated. Oncogene. 1999;18:1845–1853. - PubMed
-
- Brasier A. R., Tate J. E., Habener J. F. Optimized use of the firefly luciferase assay as a reporter gene in mammalian cell lines. Biotechniques. 1989;7:1116–1122. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
