

Evolution of the holozoan ribosome biogenesis regulon
- PMID: 18816399
- PMCID: PMC2570694
- DOI: 10.1186/1471-2164-9-442
Evolution of the holozoan ribosome biogenesis regulon
Abstract
Background: The ribosome biogenesis (RiBi) genes encode a highly-conserved eukaryotic set of nucleolar proteins involved in rRNA transcription, assembly, processing, and export from the nucleus. While the mode of regulation of this suite of genes has been studied in the yeast, Saccharomyces cerevisiae, how this gene set is coordinately regulated in the larger and more complex metazoan genomes is not understood.
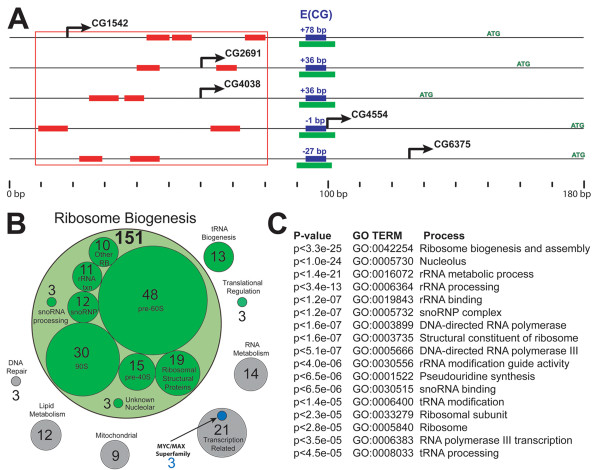
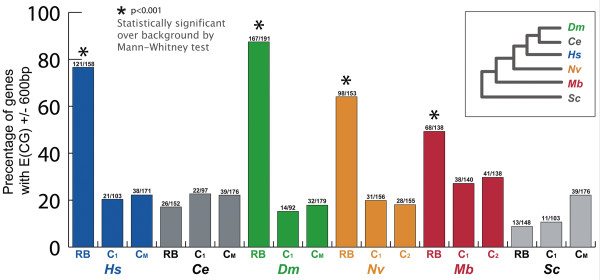
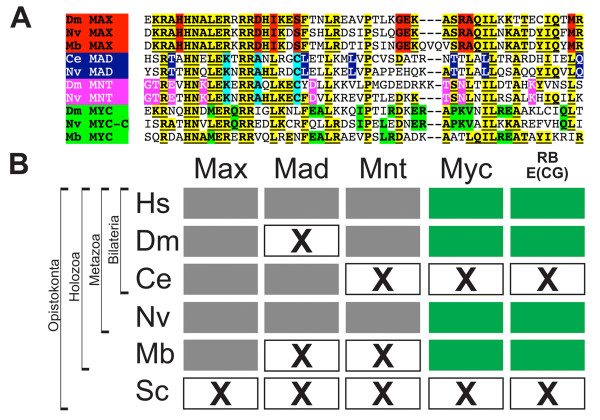
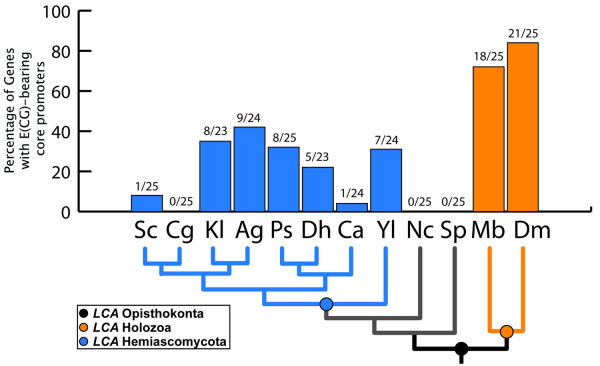
Results: Here we present genome-wide analyses indicating that a distinct mode of RiBi regulation co-evolved with the E(CG)-binding, Myc:Max bHLH heterodimer complex in a stem-holozoan, the ancestor of both Metazoa and Choanoflagellata, the protozoan group most closely related to animals. These results show that this mode of regulation, characterized by an E(CG)-bearing core-promoter, is specific to almost all of the known genes involved in ribosome biogenesis in these genomes. Interestingly, this holozoan RiBi promoter signature is absent in nematode genomes, which have not only secondarily lost Myc but are marked by invariant cell lineages typically producing small body plans of 1000 somatic cells. Furthermore, a detailed analysis of 10 fungal genomes shows that this holozoan signature in RiBi genes is not found in hemiascomycete fungi, which evolved their own unique regulatory signature for the RiBi regulon.
Conclusion: These results indicate that a Myc regulon, which is activated in proliferating cells during normal development as well as during tumor progression, has primordial roots in the evolution of an inducible growth regime in a protozoan ancestor of animals. Furthermore, by comparing divergent bHLH repertoires, we conclude that regulation by Myc but not by other bHLH genes is responsible for the evolutionary maintenance of E(CG) sites across the RiBi suite of genes.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
