

Unphosphorylated SR-like protein Npl3 stimulates RNA polymerase II elongation
- PMID: 18818768
- PMCID: PMC2538588
- DOI: 10.1371/journal.pone.0003273
Unphosphorylated SR-like protein Npl3 stimulates RNA polymerase II elongation
Abstract
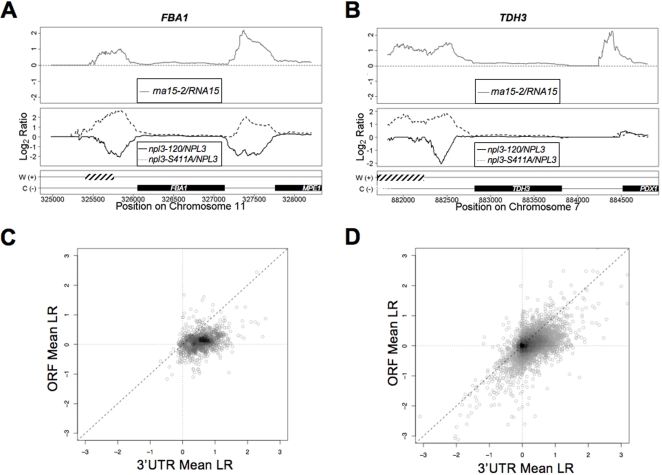
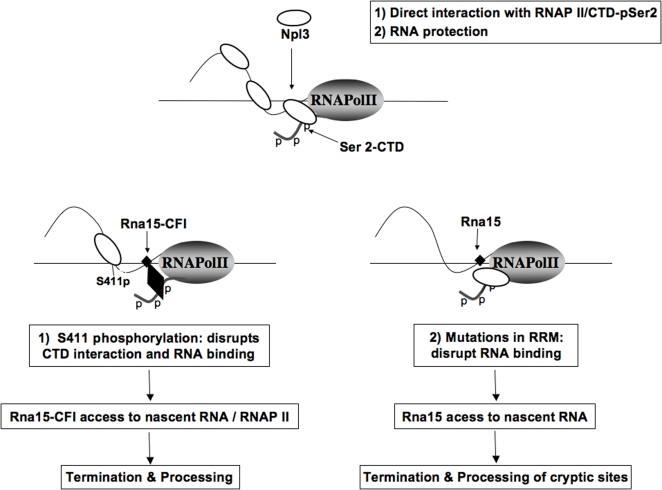
The production of a functional mRNA is regulated at every step of transcription. An area not well-understood is the transition of RNA polymerase II from elongation to termination. The S. cerevisiae SR-like protein Npl3 functions to negatively regulate transcription termination by antagonizing the binding of polyA/termination proteins to the mRNA. In this study, Npl3 is shown to interact with the CTD and have a direct stimulatory effect on the elongation activity of the polymerase. The interaction is inhibited by phosphorylation of Npl3. In addition, Casein Kinase 2 was found to be required for the phosphorylation of Npl3 and affect its ability to compete against Rna15 (Cleavage Factor I) for binding to polyA signals. Our results suggest that phosphorylation of Npl3 promotes its dissociation from the mRNA/RNAP II, and contributes to the association of the polyA/termination factor Rna15. This work defines a novel role for Npl3 in elongation and its regulation by phosphorylation.
Conflict of interest statement
Figures
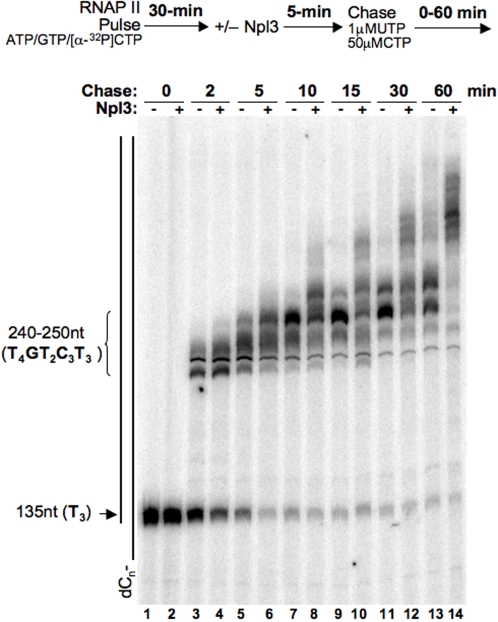
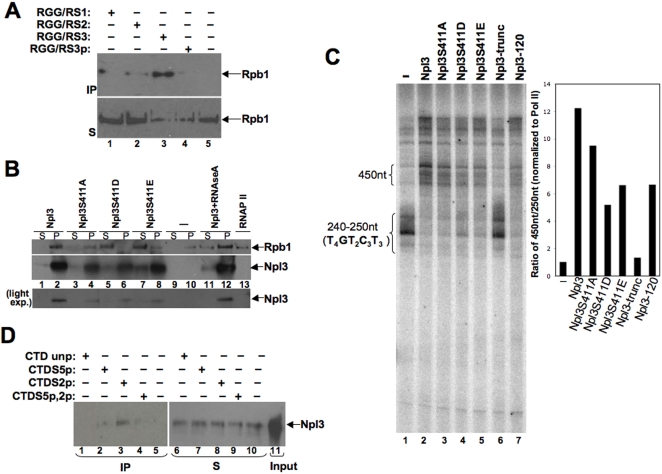
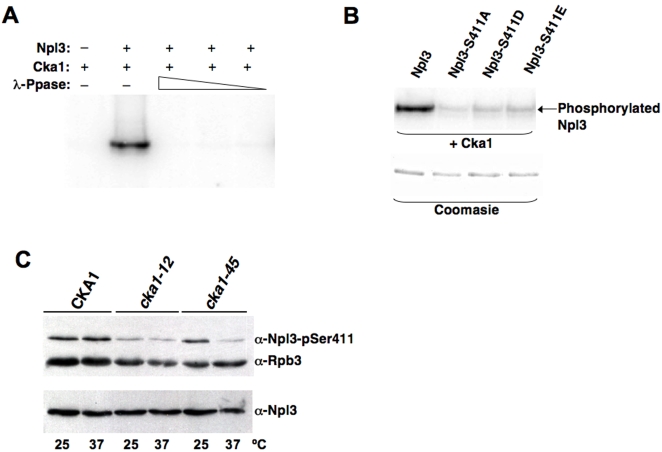
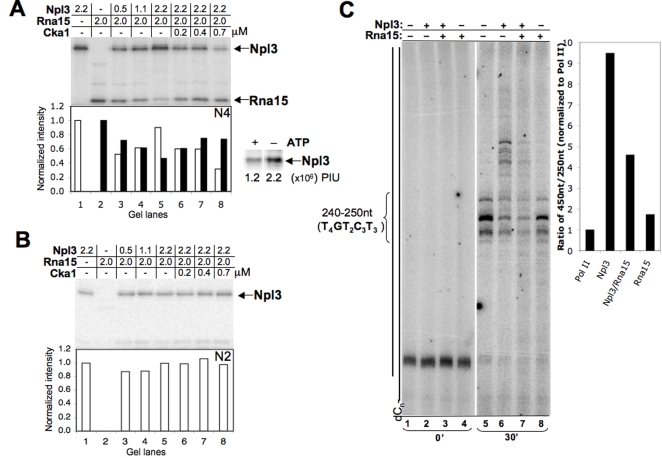
References
-
- Proudfoot NJ. How RNA polymerase II terminates transcription in higher eukaryotes. Trends Biochem Sci. 1989;14:105–110. - PubMed
-
- Hirose Y, Manley JL. RNA polymerase II and the integration of nuclear events. Genes Dev. 2000;14:1415–1429. - PubMed
-
- McCracken S, Fong N, Yankulov K, Ballantyne S, Pan G, et al. The C-terminal domain of RNA polymerase II couples mRNA processing to transcription. Nature. 1997;385:357–361. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 GM056663/GM/NIGMS NIH HHS/United States
- 5F31GM075383-03/GM/NIGMS NIH HHS/United States
- K01 CA115515/CA/NCI NIH HHS/United States
- R29 GM051124/GM/NIGMS NIH HHS/United States
- 1K01CA115515-01A1/CA/NCI NIH HHS/United States
- R01 GM068887/GM/NIGMS NIH HHS/United States
- GM56663/GM/NIGMS NIH HHS/United States
- GM51124/GM/NIGMS NIH HHS/United States
- GM68887/GM/NIGMS NIH HHS/United States
- R01 GM046498/GM/NIGMS NIH HHS/United States
- GM46498/GM/NIGMS NIH HHS/United States
- R01 GM051124/GM/NIGMS NIH HHS/United States
- F31 GM075383/GM/NIGMS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
