

doi: 10.1074/jbc.M805171200.
Epub 2008 Sep 25.
On the mechanism of pore formation by melittin
Affiliations
- PMID: 18819911
- PMCID: PMC2662212
- DOI: 10.1074/jbc.M805171200
Item in Clipboard
On the mechanism of pore formation by melittin
J Biol Chem.
.
Abstract
The mechanism of pore formation of lytic peptides, such as melittin from bee venom, is thought to involve binding to the membrane surface, followed by insertion at threshold levels of bound peptide. We show that in membranes composed of zwitterionic lipids, i.e. phosphatidylcholine, melittin not only forms pores but also inhibits pore formation. We propose that these two modes of action are the result of two competing reactions: direct insertion into the membrane and binding parallel to the membrane surface. The direct insertion of melittin leads to pore formation, whereas the parallel conformation is inactive and prevents other melittin molecules from inserting, hence preventing pore formation.
Figures
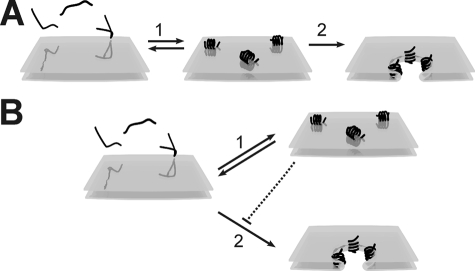
Models of pore formation by melittin in membranes composed of
zwitterionic PC lipids. A, existing model. Step 1: At
low concentrations, melittin (∼) binds to the membrane and forms an
amphipatic α-helix oriented parallel to the membrane. Step 2:
If the melittin concentration reaches above a certain threshold, melittin
inserts in the membrane and the orientation shifts to largely perpendicular,
causing pore formation. B, new model. Melittin (∼) binds and
forms an amphipatic α-helix and can be oriented either parallel
(step 1) or perpendicular (step 2) to the plane of the
membrane. The perpendicular orientation leads to membrane insertion and pore
formation, whereas the parallel orientation is inactive and prevents other
melittin molecules from forming pores, hence protecting the membrane
(dotted line).
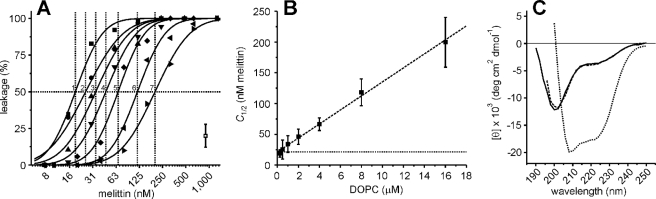
Calcein dequenching as a function of lipid (liposome) concentration.
A, calcein dequenching from liposomes composed of pure DOPC was
measured for various melittin concentrations. The final lipid concentrations
were 0.25 μm (1, ▪), 0.5 μm
(2, ○), 1 μm (3, ▴), 2
μm (4, ▾), 4 μm (5,
♦), 8 μm (6, ◂), and 16 μm
(7, ▸). Leakage took place within 5 min and was stable for over
24 h. The typical error from at least two independent experiments is
indicated. The solid lines present fits with a cumulative log normal
distribution, and these allowed us to estimate the concentration of melittin
where 50% of the calcein leaked out (C1/2, dotted
lines) (18). B,
the C1/2 values for the various lipid concentrations
obtained from A are shown. The dashed line shows the result
of the linear regression analysis, with a slope of 11 nm
melittin/μm lipids and an offset of 21 nm (dotted
line). C, CD spectra of melittin are shown. The mean residue
ellipticity [θ] is plotted as a function of the wavelength for 44
μm melittin in 10 mm potassium phosphate, pH 7, plus
various concentrations of NaCl. Melittin is in the unfolded conformation in
the absence of NaCl (solid line); 2.5 m NaCl stabilizes
the α-helical conformation of melittin (dotted line)
(25). At the NaCl
concentration used in this study (150 mm , dashed line),
melittin is predominantly in the unfolded conformation.
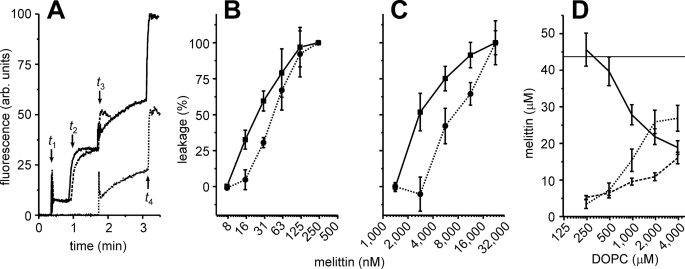
Two-step leakage experiments with melittin and DOPC liposomes.
A, calcein dequenching experiments. At time t1, 1
μm total lipid concentration in the form of liposomes were added
to the cuvette. The liposomes were loaded with either 100 mm
calcein (solid, dashed curves) or did not contain calcein
(dotted curve). At time t2, 35 nm
melittin was added to the cuvette, and leakage was determined from the changes
in fluorescence. For the solid and dotted curves, a second
batch (1 μm ) of liposomes loaded with calcein was added at time
t3. At time t4 (solid,
dotted) or time t3 (dashed), 0.03% (w/v)
of Triton X-100 was added to determine the 100% level of leakage. B,
two-step calcein dequenching experiments similar to A, with various
concentrations of melittin. The leakage of the first (solid line,
▪) and second batch of liposomes (dotted line, •) is
indicated. C, similar to B but using dual-color
fluorescence-burst analysis
(18,
20) instead of the calcein
dequenching assay, and using 250 μm lipids. The experiments from
A–C show that melittin is capable of pore formation in
freshly added liposomes but not in liposomes already present in solution.
D, reversible binding of melittin to the membranes. DOPC liposomes
were equilibrated with 44 μm of melittin, and the fractions of
liposome-bound and free melittin were determined (x-axis, lipid
concentration). The liposomes were harvested and washed by centrifugation and
then dissolved with 1% (w/v) n-dodecyl-β-d -maltoside. The
melittin concentrations of the supernatant (solid line), washing
solution (dotted line), and pellet (dashed line) were
determined by tryptophan fluorescence. Error bars are from at least 2
independent experiments.
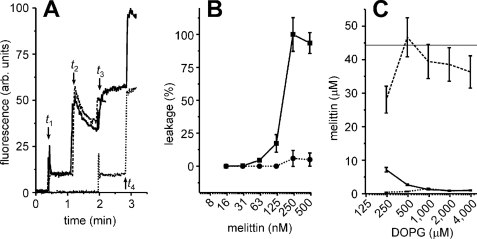
Two-step leakage experiments with melittin and DOPG liposomes.
A, calcein dequenching experiments similar to
Fig. 3A, but using 1
μm DOPG lipids in the form of liposomes and 250 nm
melittin. The decrease in fluorescence at time t2 is not
caused by photobleaching but probably by fusion of the liposomes
(18). B, two-step
calcein dequenching experiments similar to A, with various
concentrations of melittin. The leakage of the first (solid line,
▪) and second batch of liposomes (dotted line, •) is
indicated. C, same as Fig.
3D, but for liposomes composed of pure DOPG instead of
DOPC. The experiments from A–C indicate that melittin
did not cause leakage of the second batch and that all melittin irreversibly
bound to the first batch of DOPG liposomes. Error bars are from at
least two independent experiments.
Similar articles
-
The synergistic action of melittin and phospholipase A2 with lipid membranes: development of linear dichroism for membrane-insertion kinetics.Protein Pept Lett. 2010 Nov;17(11):1351-62. doi: 10.2174/0929866511009011351. Protein Pept Lett. 2010. PMID: 20673225
-
Modulation of tryptophan environment in membrane-bound melittin by negatively charged phospholipids: implications in membrane organization and function.Biochemistry. 1997 Nov 25;36(47):14291-305. doi: 10.1021/bi971933j. Biochemistry. 1997. PMID: 9398147
-
Barrel-stave model or toroidal model? A case study on melittin pores.Biophys J. 2001 Sep;81(3):1475-85. doi: 10.1016/S0006-3495(01)75802-X. Biophys J. 2001. PMID: 11509361 Free PMC article.
-
How Melittin Inserts into Cell Membrane: Conformational Changes, Inter-Peptide Cooperation, and Disturbance on the Membrane.Molecules. 2019 May 7;24(9):1775. doi: 10.3390/molecules24091775. Molecules. 2019. PMID: 31067828 Free PMC article. Review.
-
Model Membrane and Cell Studies of Antimicrobial Activity of Melittin Analogues.Curr Top Med Chem. 2016;16(1):40-5. doi: 10.2174/1568026615666150703115919. Curr Top Med Chem. 2016. PMID: 26139117 Review.
Cited by
-
Hybrid bilayer membranes on metallurgical polished aluminum.Sci Rep. 2021 May 6;11(1):9648. doi: 10.1038/s41598-021-89150-2. Sci Rep. 2021. PMID: 33958658 Free PMC article.
-
Gain-of-function analogues of the pore-forming peptide melittin selected by orthogonal high-throughput screening.J Am Chem Soc. 2012 Aug 1;134(30):12732-41. doi: 10.1021/ja3042004. Epub 2012 Jul 18. J Am Chem Soc. 2012. PMID: 22731650 Free PMC article.
-
Enhanced Therapeutic Effect of Optimized Melittin-dKLA, a Peptide Agent Targeting M2-like Tumor-Associated Macrophages in Triple-Negative Breast Cancer.Int J Mol Sci. 2022 Dec 12;23(24):15751. doi: 10.3390/ijms232415751. Int J Mol Sci. 2022. PMID: 36555393 Free PMC article.
-
Tuneable poration: host defense peptides as sequence probes for antimicrobial mechanisms.Sci Rep. 2018 Oct 8;8(1):14926. doi: 10.1038/s41598-018-33289-y. Sci Rep. 2018. PMID: 30297841 Free PMC article.
-
Reassessing the Host Defense Peptide Landscape.Front Chem. 2019 Feb 4;7:43. doi: 10.3389/fchem.2019.00043. eCollection 2019. Front Chem. 2019. PMID: 30778385 Free PMC article. Review.
References
-
- Brogden, K. A. (2005) Nat. Rev. Microbiol. 3 238–250 - PubMed
-
- Finlay, B. B., and Hancock, R. E. (2004) Nat. Rev. Microbiol. 2 497–504 - PubMed
-
- Raghuraman, H., and Chattopadhyay, A. (2007) Biosci. Rep. 27 189–223 - PubMed
-
- Son, D. J., Lee, J. W., Lee, Y. H., Song, H. S., Lee, C. K., and Hong, J. T. (2007) Pharmacol. Ther. 115 246–270 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
