

Measuring and interpreting respiratory critical oxygen pressures in roots
- PMID: 18819952
- PMCID: PMC2707311
- DOI: 10.1093/aob/mcn177
Measuring and interpreting respiratory critical oxygen pressures in roots
Abstract
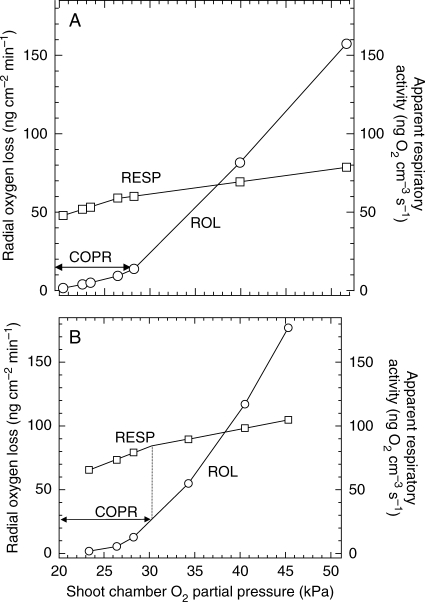
Background and aims: Respiratory critical oxygen pressures (COPR) determined from O(2)-depletion rates in media bathing intact or excised roots are unreliable indicators of respiratory O(2)-dependency in O(2)-free media and wetlands. A mathematical model was used to help illustrate this, and more relevant polarographic methods for determining COPR in roots of intact plants are discussed.
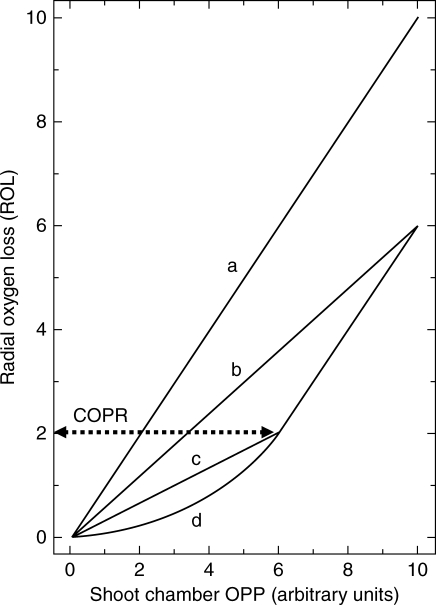
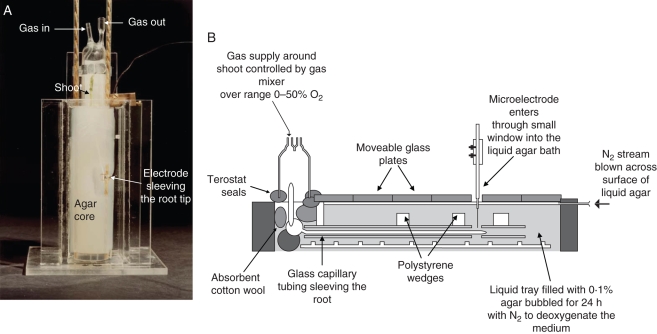
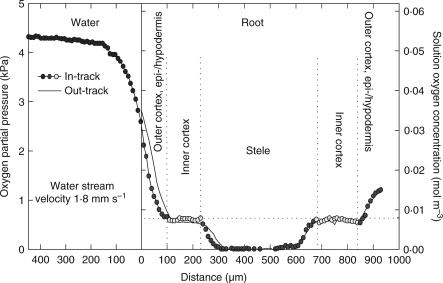
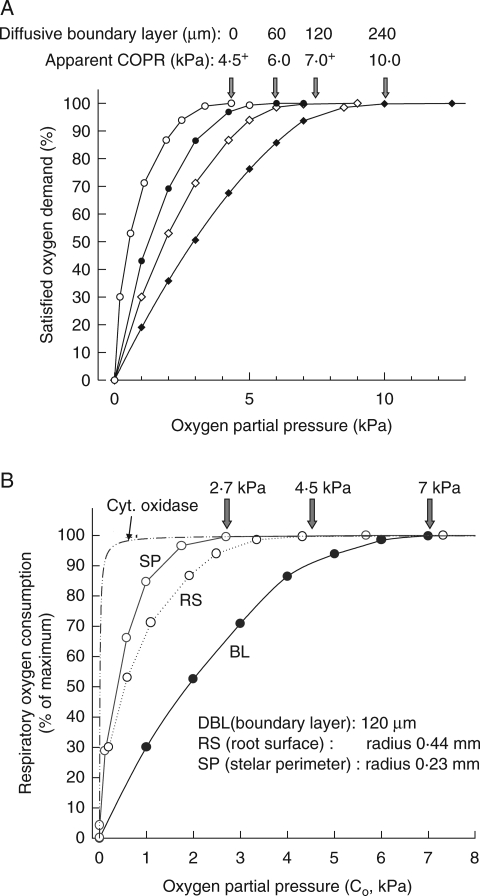
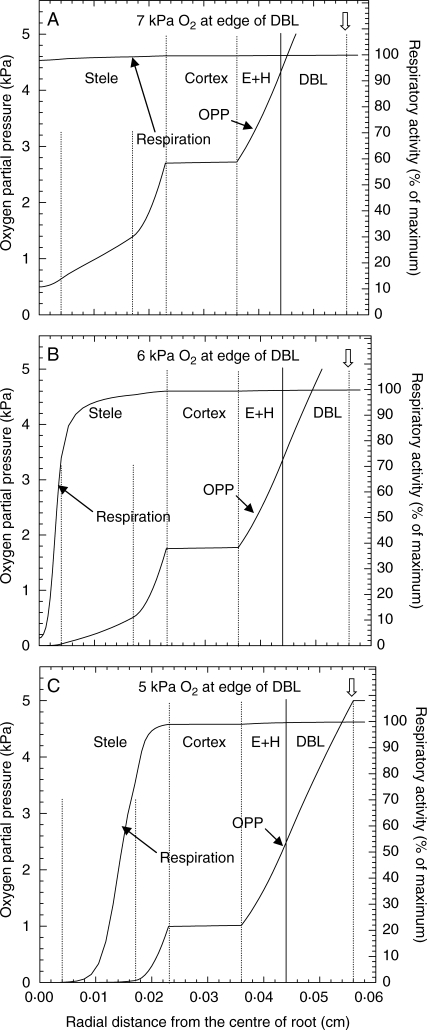
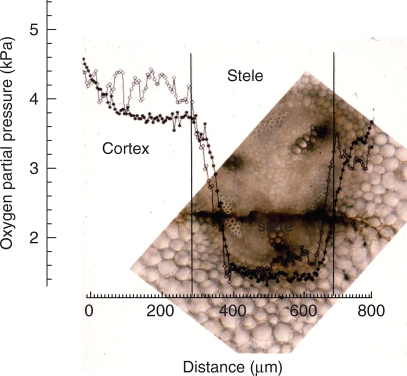
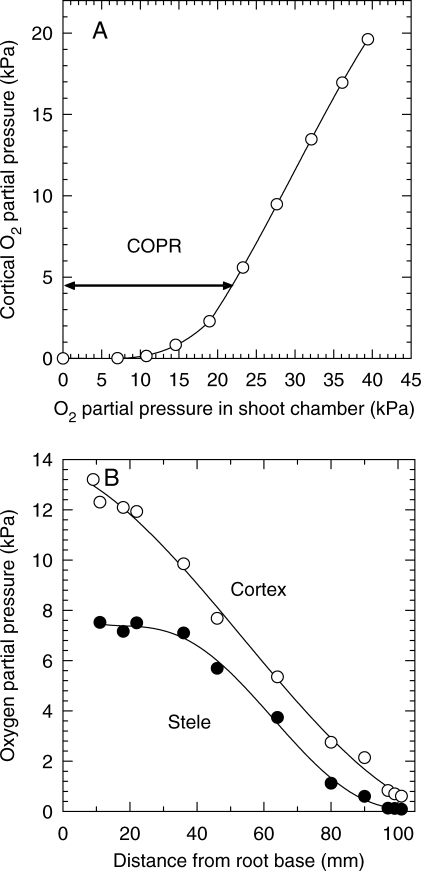
Methods: Cortical [O(2)] near the root apex was monitored indirectly (pea seedlings) from radial oxygen losses (ROL) using sleeving Pt electrodes, or directly (maize) using microelectrodes; [O(2)] in the root was controlled by manipulating [O(2)] around the shoots. Mathematical modelling of radial diffusive and respiratory properties of roots used Michaelis-Menten enzyme kinetics.
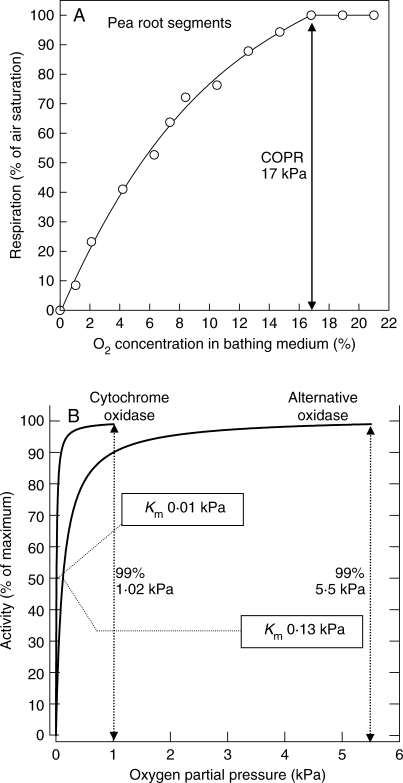
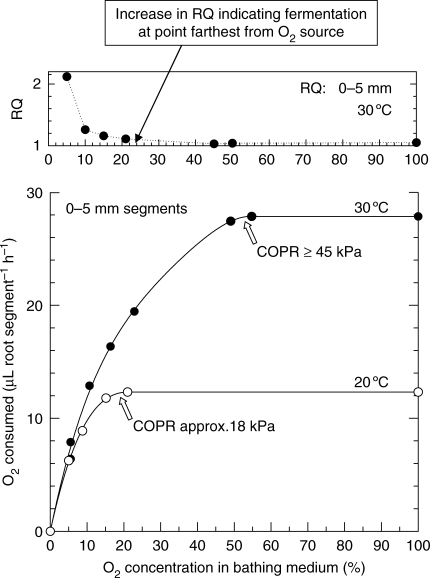
Key results: Respiration declined only when the O(2) partial pressure (OPP) in the cortex of root tips fell below 0.5-4.5 kPa, values consistent with depressed respiration near the centre of the stele as confirmed by microelectrode measurements and mathematical modelling. Modelling predictions suggested that the OPP of a significant core at the centre of roots could be below the usual detection limits of O(2)-microelectrodes but still support some aerobic respiration.
Conclusions: In O(2)-free media, as in wetlands, the COPR for roots is likely to be quite low, dependent upon the respiratory demands, dimensions and diffusion characteristics of the stele/stelar meristem and the enzyme kinetics of cytochrome oxidase. Roots of non-wetland plants may not differ greatly in their COPRs from those of wetland species. There is a possibility that trace amounts of O(2) may still be present in stelar 'anaerobic' cores where fermentation is induced at low cortical OPPs.
Figures
References
-
- Armstrong W. Aeration in higher plants. In: Woolhouse HWW, editor. Advances in botanical research. Vol. 7. New York: Academic Press; 1979. pp. 225–332.
-
- Armstrong W, Beckett PM. Root aeration in unsaturated soil: a multi-shelled model of oxygen distribution and diffusion with and without sectoral blocking of the diffusion path. New Phytologist. 1985;100:293–311.
-
- Armstrong W, Beckett PM. Internal aeration and the development of stelar anoxia in submerged roots: a multishelled mathematical model combining axial diffusion of oxygen in the cortex with radial losses to the stele, the wall layers and the rhizosphere. New Phytologist. 1987;105:221–245.
-
- Armstrong W, Drew MC. Root growth and metabolism under oxygen deficiency. In: Waisel Y, Eshel A, Kafkafi U, editors. Roots the hidden half. 3rd edn. New York: Marcel Dekker Inc; 2002. pp. 729–761.
-
- Armstrong W, Gaynard TJ. The critical oxygen pressure for root respiration in intact plants. Physiologia Plantarum. 1976;37:200–206. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
