

Mechanisms of CNS myelin inhibition: evidence for distinct and neuronal cell type specific receptor systems
- PMID: 18820405
- PMCID: PMC7259427
Mechanisms of CNS myelin inhibition: evidence for distinct and neuronal cell type specific receptor systems
Abstract
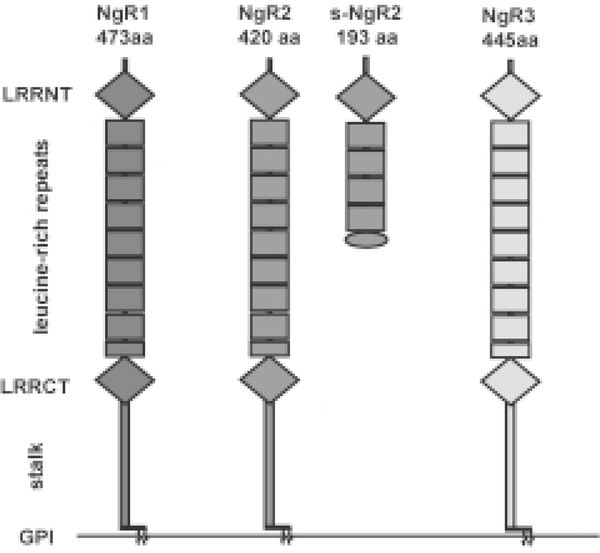
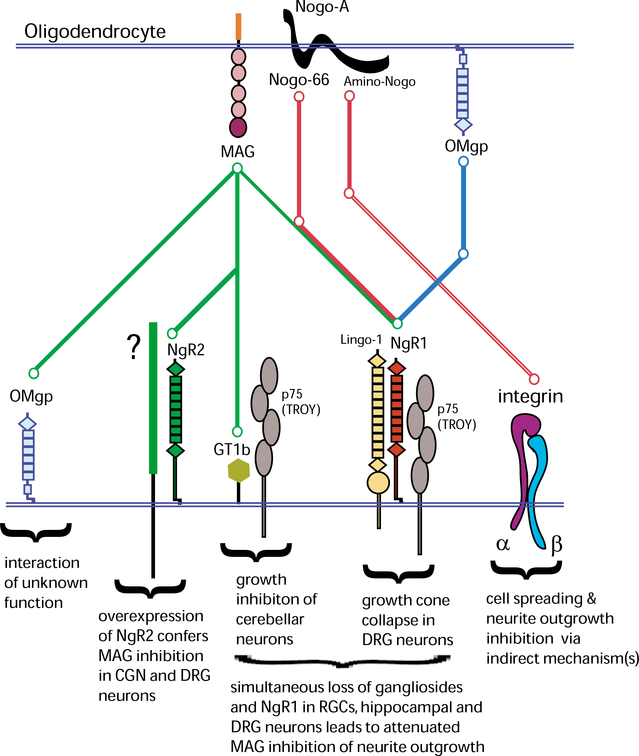
Following injury to the adult mammalian central nervous system, regenerative growth of severed axons is very limited. The lack of neuronal repair is often associated with significant functional deficits, and depending on the severity of injury, may result in permanent paralysis distal to the site of injury. A detailed understanding of the molecular mechanisms that limit neuronal growth in the injured spinal cord is an important step toward the development of specific strategies aimed at restoring functional connectivity lost as a consequence of injury. While rapid progress is being made in defining the molecular identity of CNS growth inhibitory constituents, comparatively little is known about their receptors and downstream signaling mechanisms. Emerging new evidence suggests that the mechanisms for myelin inhibition are likely to be complex, involving multiple and distinct receptor systems that may operate in a redundant manner. Furthermore, the relative contribution of a specific ligand-receptor system to bring about growth inhibition may greatly vary among different neuronal cell types. Myelin-associated glycoprotein (MAG), for example, employs different mechanisms to inhibit neurite outgrowth of cerebellar, sensory, and retinal ganglion neurons in vitro. Nogo-A harbors distinct growth inhibitory regions, which employ different signaling mechanisms. The Nogo-66 receptor 1 (NgR1), a shared ligand binding component in a receptor complex for Nogo-66, MAG, and OMgp, participates in neuronal growth cone collapse to acutely presented myelin inhibitors, but is dispensable for longitudinal neurite outgrowth inhibition on substrate-bound Nogo-66, MAG, OMgp, or crude CNS myelin in vitro. Consistent with the idea of cell-type specific mechanisms for myelin inhibition, different types of CNS neurons possess very different regenerative capacities and respond differently to experimental treatment strategies in vivo. We speculate that differences in regenerative axonal growth among different fiber systems are a reflection of their intrinsic ability to elongate axons and their distinct cell surface receptor profiles to respond to the growth inhibitory extracellular milieu. The existence of cell type specific mechanisms to impair regenerative axonal growth in the CNS may have important implications for the development of treatment strategies. Depending on the fiber tract injured, different ligand-receptor systems may need to be targeted in order to elicit robust and long-distance regenerative axonal growth.
Figures
References
-
- Barrette B, Vallieres N, Dube M, & Lacroix S (2007). Expression profile of receptors for myelin-associated inhibitors of axonal regeneration in the intact and injured mouse central nervous system. Mol Cell Neurosci, 34, 519–538. - PubMed
-
- Bartsch U, Bandtlow CE, Schnell L, Bartsch S, Spillmann AA, Rubin BP, et al. (1995). Lack of evidence that myelin-associated glycoprotein is a major inhibitor of axonal regeneration in the CNS. Neuron, 15, 1375–1381. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials