

An acceptor-substrate binding site determining glycosyl transfer emerges from mutant analysis of a plant vacuolar invertase and a fructosyltransferase
- PMID: 18821058
- PMCID: PMC2709226
- DOI: 10.1007/s11103-008-9404-7
An acceptor-substrate binding site determining glycosyl transfer emerges from mutant analysis of a plant vacuolar invertase and a fructosyltransferase
Abstract
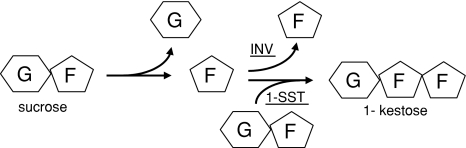
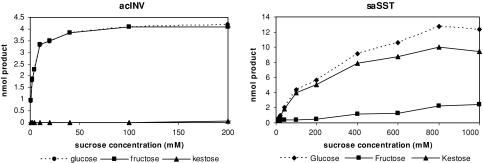
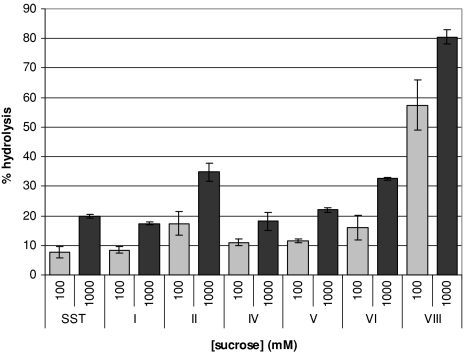
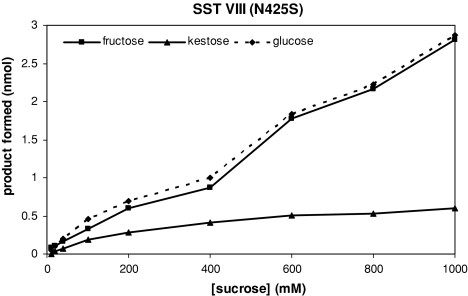
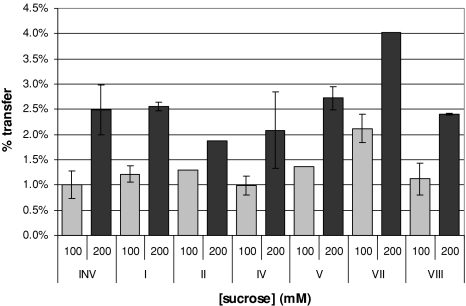
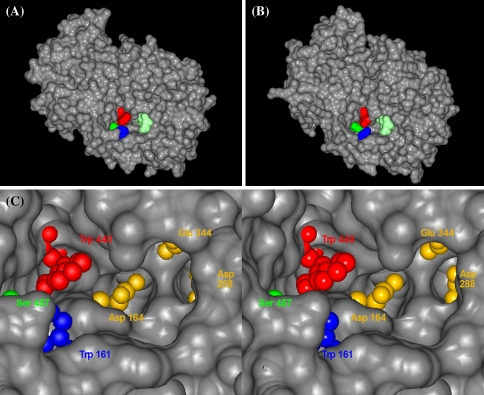
Glycoside hydrolase family 32 (GH32) harbors hydrolyzing and transglycosylating enzymes that are highly homologous in their primary structure. Eight amino acids dispersed along the sequence correlated with either hydrolase or glycosyltransferase activity. These were mutated in onion vacuolar invertase (acINV) according to the residue in festuca sucrose:sucrose 1-fructosyltransferase (saSST) and vice versa. acINV(W440Y) doubles transferase capacity. Reciprocally, saSST(C223N) and saSST(F362Y) double hydrolysis. SaSST(N425S) shows a hydrolyzing activity three to four times its transferase activity. Interestingly, modeling acINV and saSST according to the 3D structure of crystallized GH32 enzymes indicates that mutations saSST(N425S), acINV(W440Y), and the previously reported acINV(W161Y) reside very close together at the surface in the entrance of the active-site pocket. Residues in- and outside the sucrose-binding box determine hydrolase and transferase capabilities of GH32 enzymes. Modeling suggests that residues dispersed along the sequence identify a location for acceptor-substrate binding in the 3D structure of fructosyltransferases.
Figures
References
-
- {'text': '', 'ref_index': 1, 'ids': [{'type': 'DOI', 'value': '10.1074/jbc.M313911200', 'is_inner': False, 'url': 'https://doi.org/10.1074/jbc.m313911200'}, {'type': 'PubMed', 'value': '14973124', 'is_inner': True, 'url': 'https://pubmed.ncbi.nlm.nih.gov/14973124/'}]}
- Alberto F, Bignon C, Sulzenbacher G, Henrissat B, Czjzek M (2004) The three-dimensional structure of invertase (β-fructosidase) from Thermotoga maritima reveals a bimodular arrangement and an evolutionary relationship between retaining and inverting glycosidases. J Biol Chem 279:18903–18910. doi:10.1074/jbc.M313911200 - PubMed
-
- {'text': '', 'ref_index': 1, 'ids': [{'type': 'DOI', 'value': '10.1016/j.febslet.2004.04.064', 'is_inner': False, 'url': 'https://doi.org/10.1016/j.febslet.2004.04.064'}, {'type': 'PubMed', 'value': '15178325', 'is_inner': True, 'url': 'https://pubmed.ncbi.nlm.nih.gov/15178325/'}]}
- Altenbach D, Nüesch E, Meyer AD, Boller T, Wiemken A (2004) The large subunit determines catalytic specificity of barley sucrose:fructan 6-fructosyltransferase and fescue sucrose:sucrose 1-fructosyltransferase. FEBS Lett 567:214–218. doi:10.1016/j.febslet.2004.04.064 - PubMed
-
- {'text': '', 'ref_index': 1, 'ids': [{'type': 'DOI', 'value': '10.1016/j.febslet.2005.07.034', 'is_inner': False, 'url': 'https://doi.org/10.1016/j.febslet.2005.07.034'}, {'type': 'PubMed', 'value': '16098522', 'is_inner': True, 'url': 'https://pubmed.ncbi.nlm.nih.gov/16098522/'}]}
- Altenbach D, Nüesch E, Ritsema T, Boller T, Wiemken A (2005) Mutational analysis of the active center of plant fructosyltransferases: Festuca 1-SST and barley 6-SFT. FEBS Lett 579:4647–4653. doi:10.1016/j.febslet.2005.07.034 - PubMed
-
- {'text': '', 'ref_index': 1, 'ids': [{'type': 'DOI', 'value': '10.1107/S0907444998003254', 'is_inner': False, 'url': 'https://doi.org/10.1107/s0907444998003254'}, {'type': 'PubMed', 'value': '9757107', 'is_inner': True, 'url': 'https://pubmed.ncbi.nlm.nih.gov/9757107/'}]}
- Brünger AT, Adams PD, Clore GM, DeLano WL, Gros P, Grosse-Kunstleve RW et al (1998) Crystallography & NMR system: a new software suite for macromolecular structure determination. Acta Crystallogr D Biol Crystallogr 54:905–921. doi:10.1107/S0907444998003254 - PubMed
-
- {'text': '', 'ref_index': 1, 'ids': [{'type': 'DOI', 'value': '10.1111/j.1469-8137.1989.tb00322.x', 'is_inner': False, 'url': 'https://doi.org/10.1111/j.1469-8137.1989.tb00322.x'}]}
- Cairns AJ, Winters A, Pollock CJ (1989) Fructan biosynthesis in excised leaves of Lolium temulentum L.III. A comparison of the in vitro properties of fructosyl transferase activities with the characteristics of in vivo fructan accumulation. New Phytol 112:343–352. doi:10.1111/j.1469-8137.1989.tb00322.x
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
