

Protective effect of Pycnogenol in human neuroblastoma SH-SY5Y cells following acrolein-induced cytotoxicity
- PMID: 18822368
- PMCID: PMC2849727
- DOI: 10.1016/j.freeradbiomed.2008.08.025
Protective effect of Pycnogenol in human neuroblastoma SH-SY5Y cells following acrolein-induced cytotoxicity
Abstract
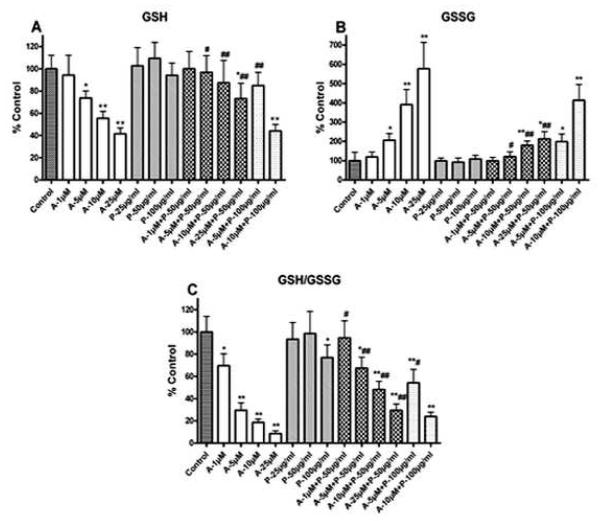
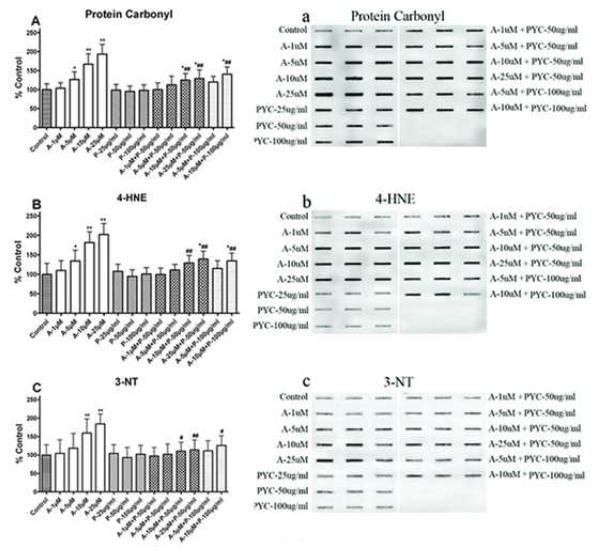
Oxidative stress is one of the hypotheses involved in the etiology of Alzheimer's disease (AD). Considerable attention has been focused on increasing the intracellular glutathione (GSH) levels in many neurodegenerative diseases, including AD. Pycnogenol (PYC) has antioxidant properties and stabilizes intracellular antioxidant defense systems including glutathione levels. The present study investigated the protective effects of PYC on acrolein-induced oxidative cell toxicity in cultured SH-SY5Y neuroblastoma cells. Decreased cell survival in SH-SY5Y cultures treated with acrolein correlated with oxidative stress, increased NADPH oxidase activity, free radical production, protein oxidation/nitration (protein carbonyl, 3-nitrotyrosine), and lipid peroxidation (4-hydroxy-2-nonenal). Pretreatment with PYC significantly attenuated acrolein-induced cytotoxicity, protein damage, lipid peroxidation, and cell death. A dose-response study suggested that PYC showed protective effects against acrolein toxicity by modulating oxidative stress and increasing GSH. These findings provide support that PYC may provide a promising approach for the treatment of oxidative stress-related neurodegenerative diseases such as AD.
Figures







References
-
- Kondo M, Oya-Ito T, Kumagai T, Osawa T, Uchida K. Cyclopentenone prostaglandins as potential inducers of intracellular oxidative stress. The Journal of biological chemistry. 2001;276:12076–12083. - PubMed
-
- Naoi M, Maruyama W, Shamoto-Nagai M, Yi H, Akao Y, Tanaka M. Oxidative stress in mitochondria: decision to survival and death of neurons in neurodegenerative disorders. Molecular neurobiology. 2005;31:81–93. - PubMed
-
- Sundaresan M, Yu ZX, Ferrans VJ, Irani K, Finkel T. Requirement for generation of H2O2 for platelet-derived growth factor signal transduction. Science (New York, N.Y. 1995;270:296–299. - PubMed
-
- Ansari MA, Joshi G, Huang Q, Opii WO, Abdul HM, Sultana R, Butterfield DA. In vivo administration of D609 leads to protection of subsequently isolated gerbil brain mitochondria subjected to in vitro oxidative stress induced by amyloid beta-peptide and other oxidative stressors: relevance to Alzheimer’s disease and other oxidative stress-related neurodegenerative disorders. Free radical biology & medicine. 2006;41:1694–1703. - PMC - PubMed
-
- Greenamyre JT, Betarbet R, Sherer TB. The rotenone model of Parkinson’s disease: genes, environment and mitochondria. Parkinsonism & related disorders. 2003;9(Suppl 2):S59–64. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
