

Leishmania donovani depletes labile iron pool to exploit iron uptake capacity of macrophage for its intracellular growth
- PMID: 18823384
- PMCID: PMC2774478
- DOI: 10.1111/j.1462-5822.2008.01241.x
Leishmania donovani depletes labile iron pool to exploit iron uptake capacity of macrophage for its intracellular growth
Abstract
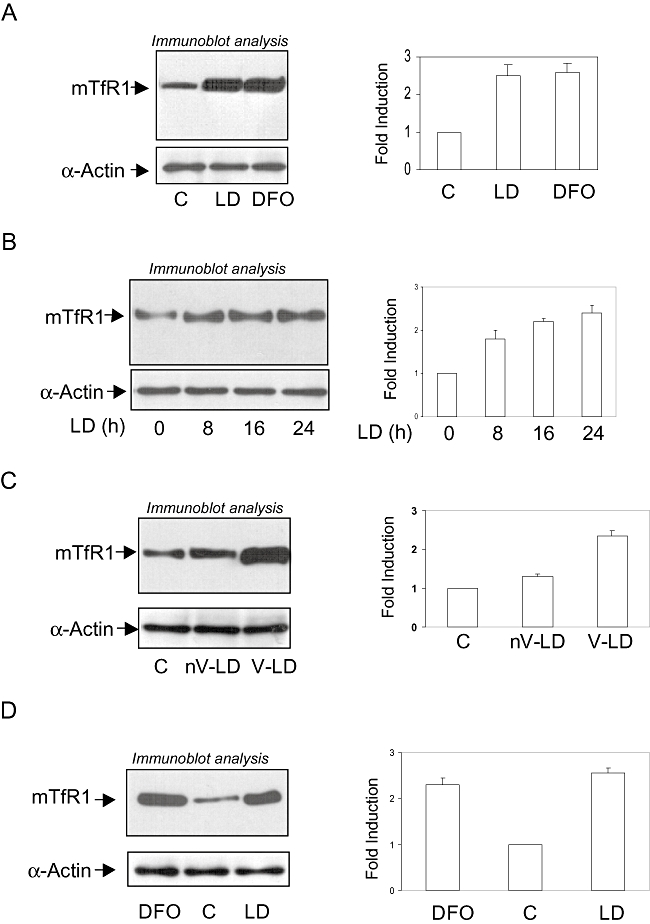
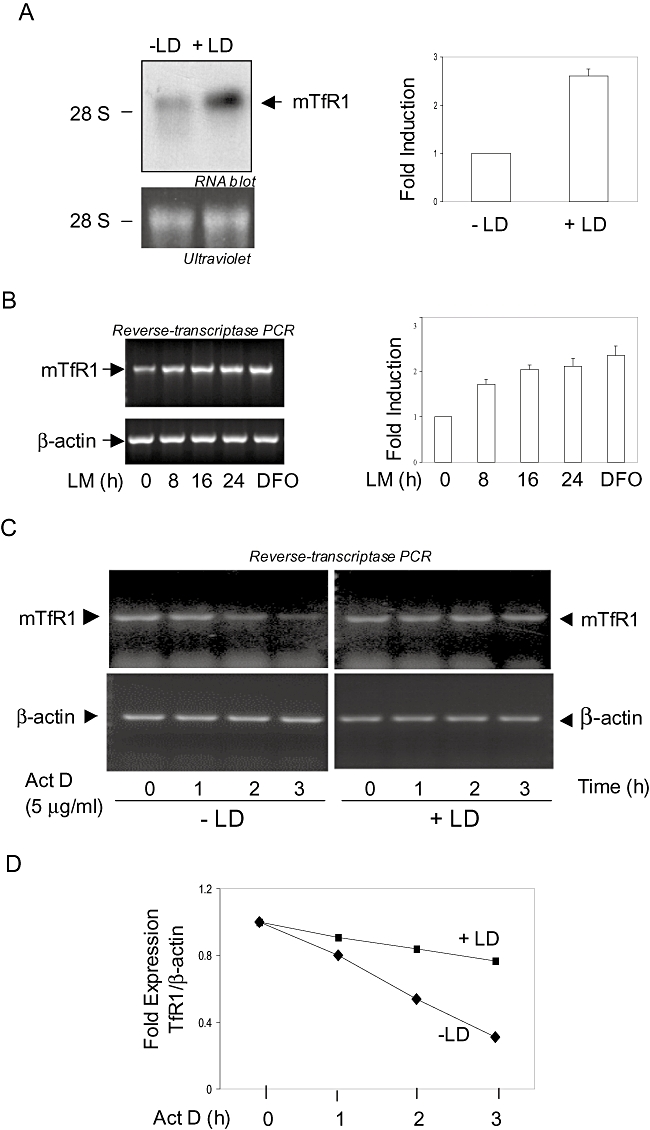
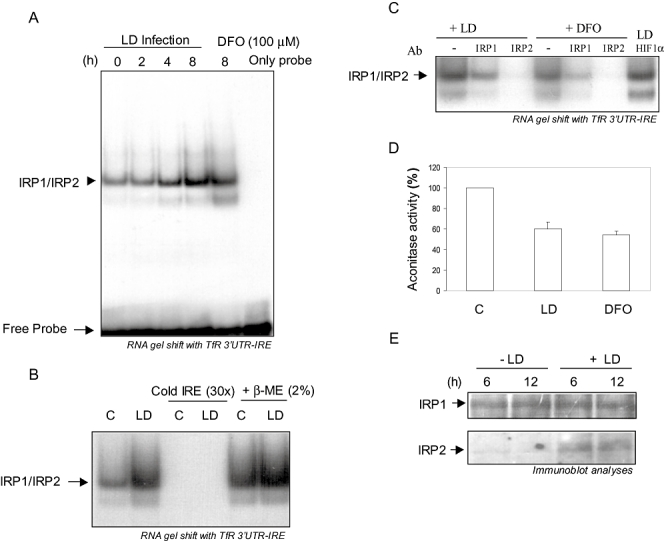
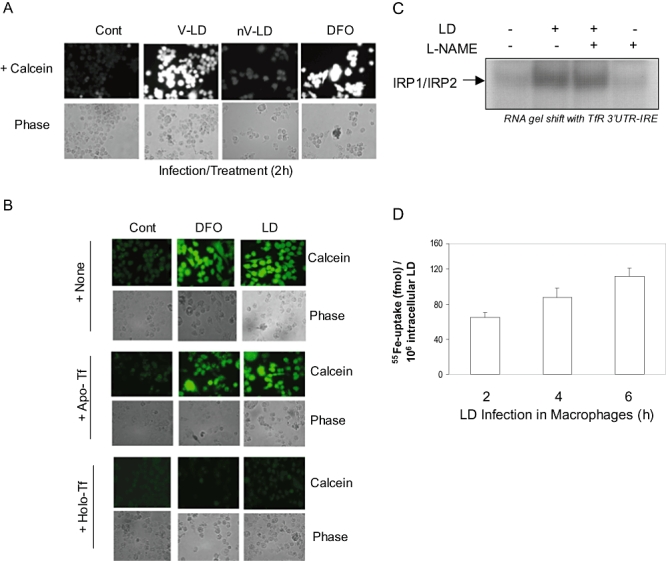
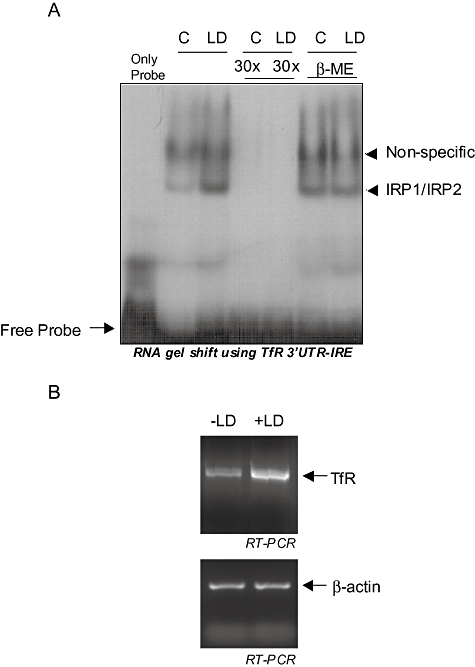
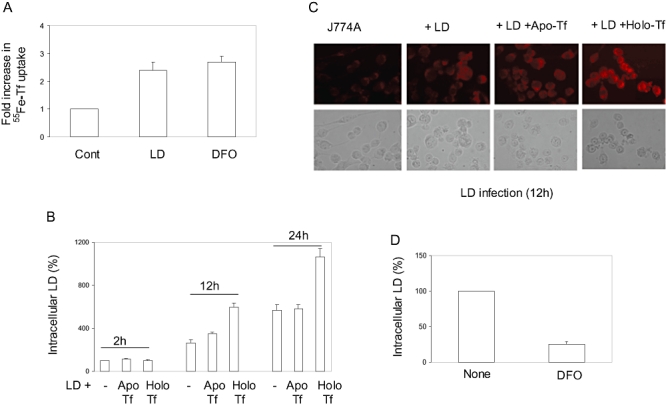
Intracellular pathogens employ several strategies for iron acquisition from host macrophages for survival and growth, whereas macrophage resists infection by actively sequestering iron. Here, we show that instead of allowing macrophage to sequester iron, protozoan parasite Leishmania donovani (LD) uses a novel strategy to manipulate iron uptake mechanisms of the host and utilizes the taken up iron for its intracellular growth. To do so, intracellular LD directly scavenges iron from labile iron pool of macrophages. Depleted labile iron pool activates iron sensors iron-regulatory proteins IRP1 and IRP2. IRPs then bind to iron-responsive elements present in the 3' UTR of iron uptake gene transferrin receptor 1 by a post-transcriptional mRNA stability mechanism. Increased iron-responsive element-IRP interaction and transferrin receptor 1 expressions in spleen-derived macrophages from LD-infected mice confirm that LD employs similar mechanism to acquire iron during infection into mammalian hosts. Increased intracellular LD growth by holo-transferrin supplementation and inhibited growth by iron chelator treatment confirm the significance of this modulated iron uptake pathway of host in favour of the parasite.
Figures
Similar articles
-
Leishmania donovani inhibits ferroportin translation by modulating FBXL5-IRP2 axis for its growth within host macrophages.Cell Microbiol. 2018 Jul;20(7):e12834. doi: 10.1111/cmi.12834. Epub 2018 Mar 30. Cell Microbiol. 2018. PMID: 29470856
-
Iron regulatory protein 1 outcompetes iron regulatory protein 2 in regulating cellular iron homeostasis in response to nitric oxide.J Biol Chem. 2011 Jul 1;286(26):22846-54. doi: 10.1074/jbc.M111.231902. Epub 2011 May 12. J Biol Chem. 2011. PMID: 21566147 Free PMC article.
-
Excess capacity of the iron regulatory protein system.J Biol Chem. 2007 Aug 24;282(34):24650-9. doi: 10.1074/jbc.M703167200. Epub 2007 Jun 28. J Biol Chem. 2007. PMID: 17604281
-
Mammalian iron metabolism and its control by iron regulatory proteins.Biochim Biophys Acta. 2012 Sep;1823(9):1468-83. doi: 10.1016/j.bbamcr.2012.05.010. Epub 2012 May 17. Biochim Biophys Acta. 2012. PMID: 22610083 Free PMC article. Review.
-
Role of nitric oxide in cellular iron metabolism.Biometals. 2003 Mar;16(1):125-35. doi: 10.1023/a:1020788603046. Biometals. 2003. PMID: 12572672 Review.
Cited by
-
Iron in intracellular infection: to provide or to deprive?Front Cell Infect Microbiol. 2013 Dec 9;3:96. doi: 10.3389/fcimb.2013.00096. eCollection 2013. Front Cell Infect Microbiol. 2013. PMID: 24367768 Free PMC article. Review.
-
Leishmania-mediated inhibition of iron export promotes parasite replication in macrophages.PLoS Pathog. 2014 Jan 30;10(1):e1003901. doi: 10.1371/journal.ppat.1003901. eCollection 2014 Jan. PLoS Pathog. 2014. PMID: 24497831 Free PMC article.
-
Genetic Iron Overload Hampers Development of Cutaneous Leishmaniasis in Mice.Int J Mol Sci. 2023 Jan 14;24(2):1669. doi: 10.3390/ijms24021669. Int J Mol Sci. 2023. PMID: 36675185 Free PMC article.
-
Chlamydia trachomatis alters iron-regulatory protein-1 binding capacity and modulates cellular iron homeostasis in HeLa-229 cells.J Biomed Biotechnol. 2009;2009:342032. doi: 10.1155/2009/342032. Epub 2009 Aug 16. J Biomed Biotechnol. 2009. PMID: 19688112 Free PMC article.
-
Oral activity of the antimalarial endoperoxide 6-(1,2,6,7-tetraoxaspiro[7.11]nonadec-4-yl)hexan-1-ol (N-251) against Leishmania donovani complex.PLoS Negl Trop Dis. 2019 Mar 25;13(3):e0007235. doi: 10.1371/journal.pntd.0007235. eCollection 2019 Mar. PLoS Negl Trop Dis. 2019. PMID: 30908481 Free PMC article.
References
-
- Andrews NC. Probing the iron pool. Focus on ‘Detection of intracellular iron by its regulatory effect’. Am J Physiol Cell Physiol. 2004;287:C1537–C1538. - PubMed
-
- Andrews NC, Schmidt PJ. Iron homeostasis. Annu Rev Physiol. 2007;69:69–85. - PubMed
-
- Borges VM, Vannier-Santos MA, de Souza W. Subverted transferrin trafficking in Leishmania-infected macrophages. Parasitol Res. 1998;84:811–822. - PubMed
-
- Braun V, Killmann H. Bacterial solutions to the iron-supply problem. Trends Biochem Sci. 1999;24:104–109. - PubMed
-
- Chang KP. Human cutaneous lieshmania in a mouse macrophage line: propagation and isolation of intracellular parasites. Science. 1980;209:1240–1242. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
