

Rhythm generation by the pre-Bötzinger complex in medullary slice and island preparations: effects of adenosine A(1) receptor activation
- PMID: 18826652
- PMCID: PMC2567986
- DOI: 10.1186/1471-2202-9-95
Rhythm generation by the pre-Bötzinger complex in medullary slice and island preparations: effects of adenosine A(1) receptor activation
Abstract
Background: The pre-Bötzinger complex (preBötC) is a central pattern generator within the ventrolateral medulla oblongata's ventral respiratory group that is important for the generation of respiratory rhythm. Activation of adenosine A(1) receptors (A(1)R) depresses preBötC rhythmogenesis. Although it remains unclear whether A(1)R activation is important for organisms in a normal metabolic state, A(1)R activation is important to the response of the preBötC to metabolic stress, such as hypoxia. This study examined mechanisms linking A(1)R activation to depression of preBötC rhythmogenesis in medullary slice and island preparations from neonatal mice.
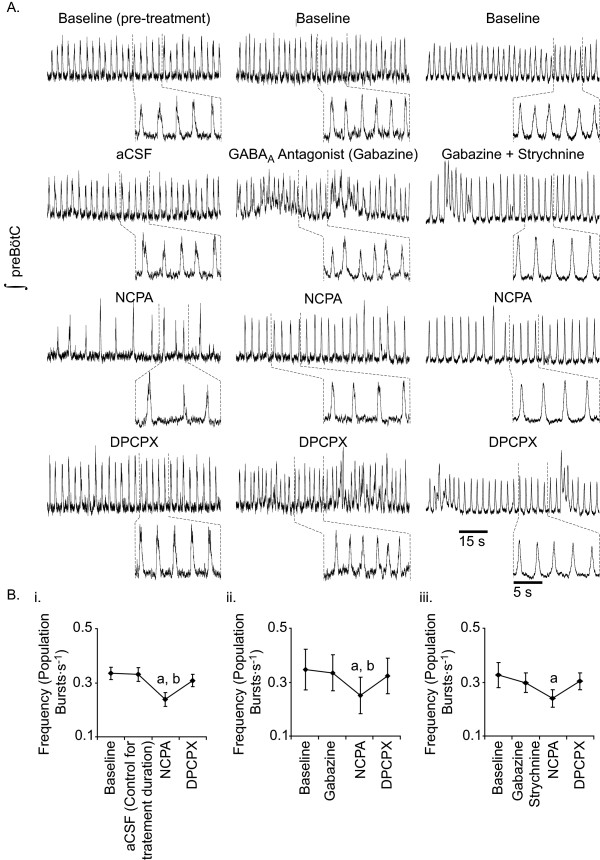
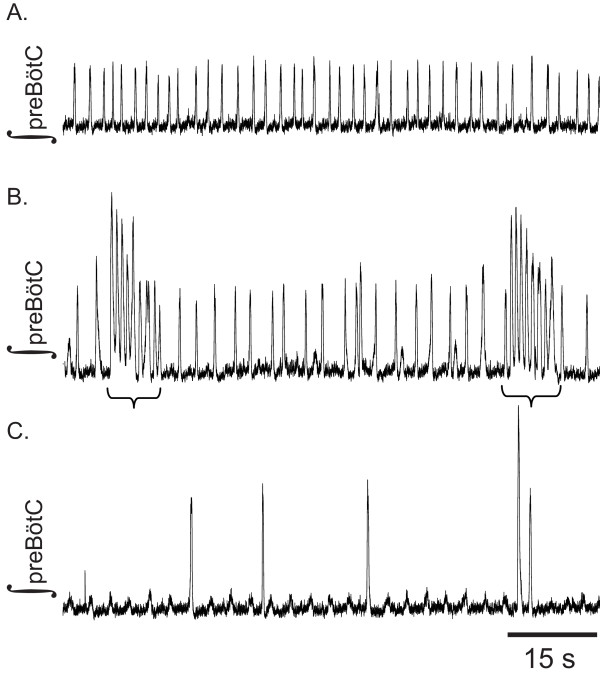
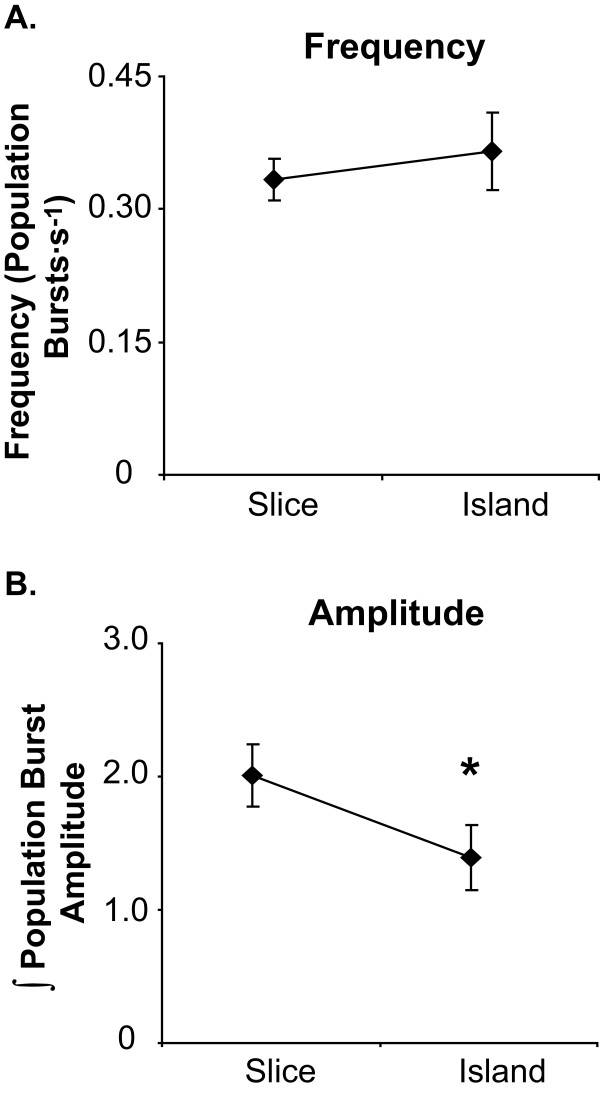
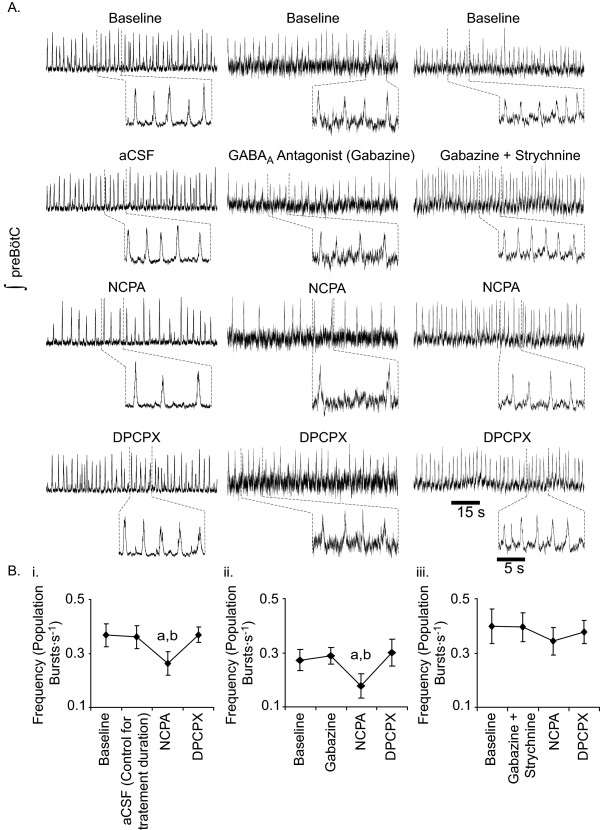
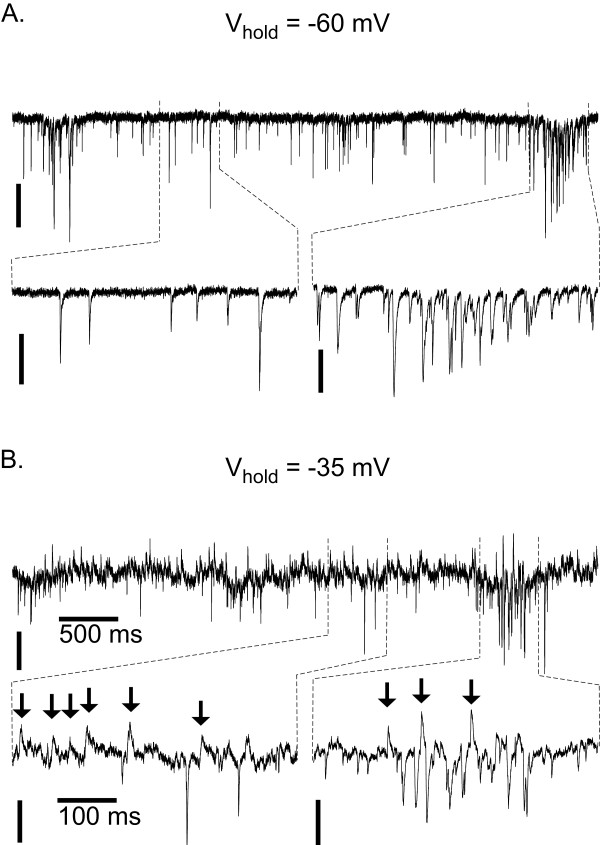
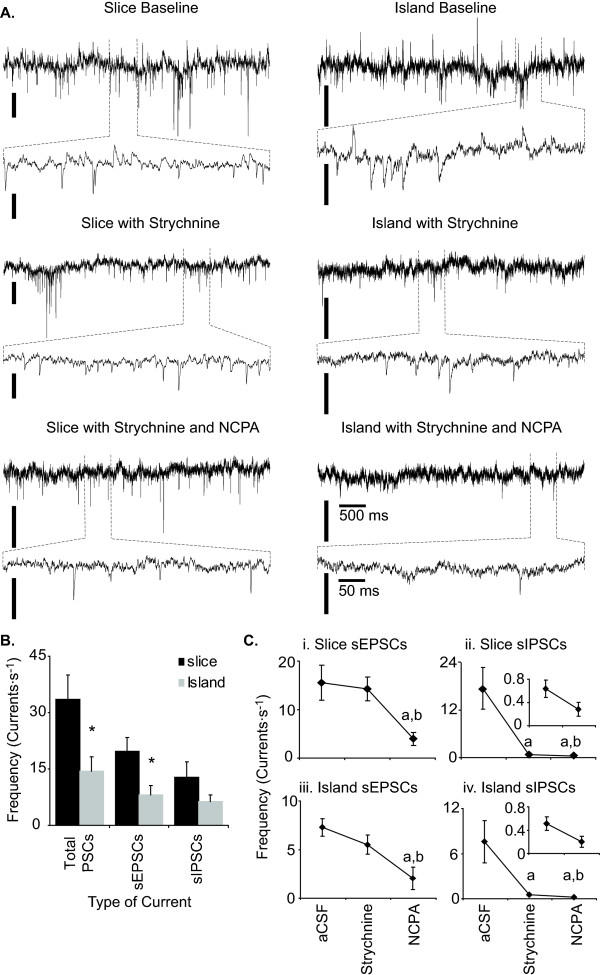
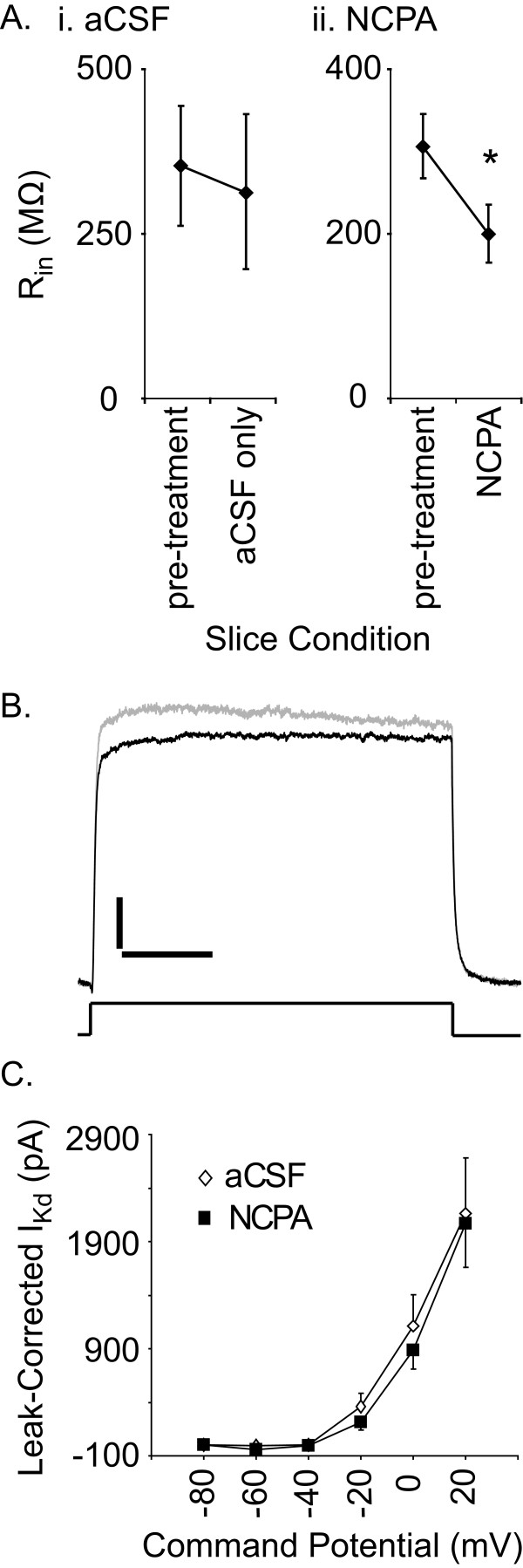
Results: Converting medullary slices to islands by cutting away much of the medullary tissue adjacent to the preBötC decreased the amplitude of action potential bursts generated by a population of neurons within the preBötC (recorded with an extracellular electrode, and integrated using a hardware integrator), without noticeably affecting burst frequency. The A(1)R agonist N6-Cyclopentyladenosine (NCPA) reduced population burst frequency in slices by ca. 33% and in islands by ca. 30%. As in normal (drug-free) artificial cerebrospinal fluid (aCSF), NCPA decreased burst frequency in slices when GABA(A)ergic or GABA(A)ergic and glycinergic transmission were blocked, and in islands when GABA(A)ergic transmission was antagonized. Converting slices to island preparations decreased synaptic input to inspiratory neurons. NCPA further decreased the frequency of synaptic inputs to neurons in island preparations and lowered the input resistance of inspiratory neurons, even when chemical communication between neurons and other cells was impeded.
Conclusion: Together these data support the suggestion that depression of preBötC activity by A(1)R activation involves both decreased neuronal excitability and diminished inter-neuronal communication.
Figures
References
-
- Thoby-Brisson M, Ramirez JM. Identification of two types of inspiratory pacemaker neurons in the isolated respiratory neural network of mice. J Neurophysiol. 2001;86:104–112. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
