

Promoter DNA hypermethylation and gene repression in undifferentiated Arabidopsis cells
- PMID: 18827894
- PMCID: PMC2556100
- DOI: 10.1371/journal.pone.0003306
Promoter DNA hypermethylation and gene repression in undifferentiated Arabidopsis cells
Abstract
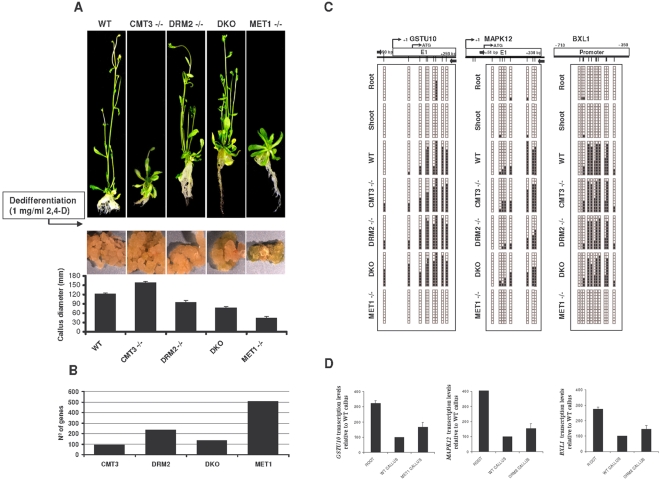
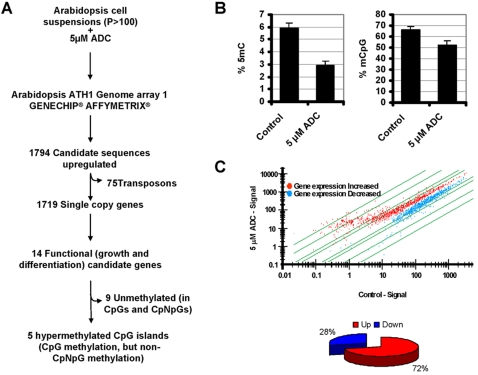
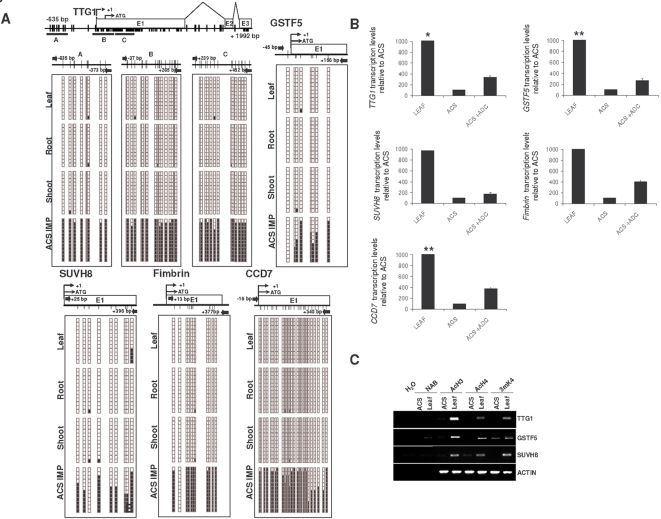
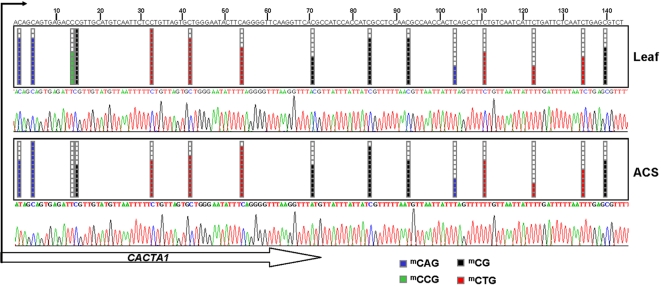
Maintaining and acquiring the pluripotent cell state in plants is critical to tissue regeneration and vegetative multiplication. Histone-based epigenetic mechanisms are important for regulating this undifferentiated state. Here we report the use of genetic and pharmacological experimental approaches to show that Arabidopsis cell suspensions and calluses specifically repress some genes as a result of promoter DNA hypermethylation. We found that promoters of the MAPK12, GSTU10 and BXL1 genes become hypermethylated in callus cells and that hypermethylation also affects the TTG1, GSTF5, SUVH8, fimbrin and CCD7 genes in cell suspensions. Promoter hypermethylation in undifferentiated cells was associated with histone hypoacetylation and primarily occurred at CpG sites. Accordingly, we found that the process specifically depends on MET1 and DRM2 methyltransferases, as demonstrated with DNA methyltransferase mutants. Our results suggest that promoter DNA methylation may be another important epigenetic mechanism for the establishment and/or maintenance of the undifferentiated state in plant cells.
Conflict of interest statement
Figures

References
-
- Grafi G. How cells dedifferentiate: a lesson from plants. Dev Biol. 2004;268:1–6. - PubMed
-
- Doerner P. Plant meristems: what you see is what you get? Curr Biol. 2006;16:R56–58. - PubMed
-
- Grafi G, Avivi Y. Stem cells: a lesson from dedifferentiation. Trends Biotechnol. 2004;22:388–389. - PubMed
-
- Costa S, Shaw P. ‘Open minded’ cells: how cells can change fate. Trends Cell Biol. 2007;17:101–106. - PubMed
-
- Lippman Z, Gendrel AV, Black M, Vaughn MW, Dedhia N, et al. Role of transposable elements in heterochromatin and epigenetic control. Nature. 2004;430:471–476. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
