

Social modulation during songbird courtship potentiates midbrain dopaminergic neurons
- PMID: 18827927
- PMCID: PMC2533700
- DOI: 10.1371/journal.pone.0003281
Social modulation during songbird courtship potentiates midbrain dopaminergic neurons
Abstract
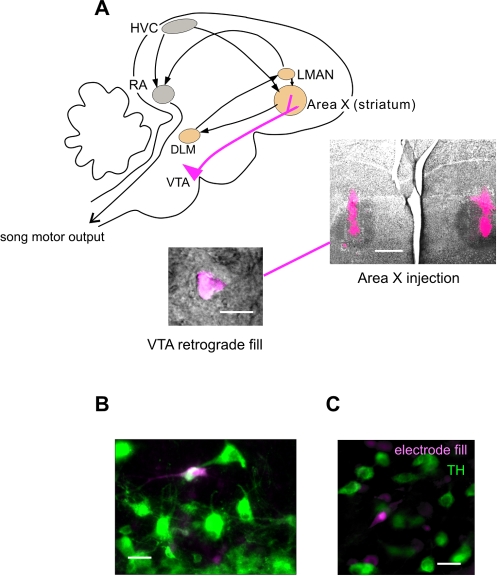
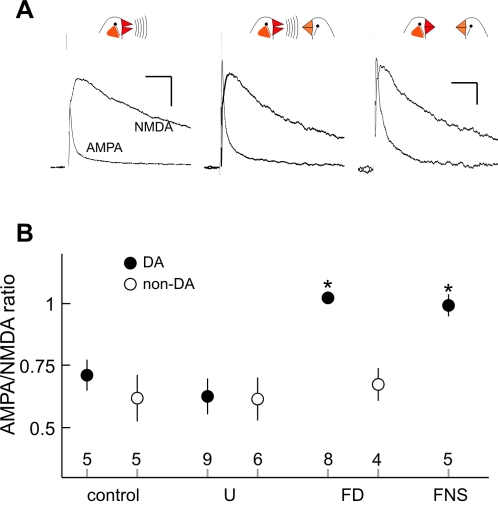
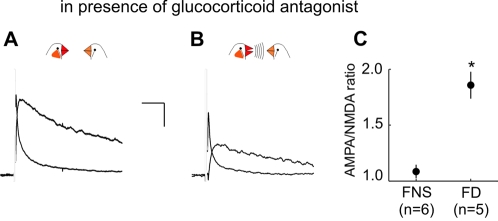
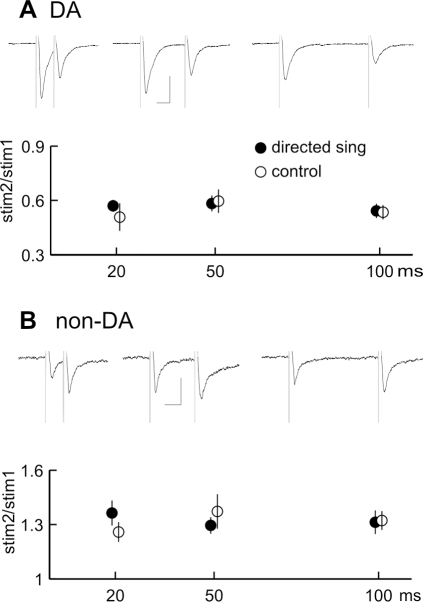
Synaptic transmission onto dopaminergic neurons of the mammalian ventral tegmental area (VTA) can be potentiated by acute or chronic exposure to addictive drugs. Because rewarding behavior, such as social affiliation, can activate the same neural circuitry as addictive drugs, we tested whether the intense social interaction of songbird courtship may also potentiate VTA synaptic function. We recorded glutamatergic synaptic currents from VTA of male zebra finches who had experienced distinct social and behavioral conditions during the previous hour. The level of synaptic transmission to VTA neurons, as assayed by the ratio of alpha-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA) to N-methyl-D-aspartic acid (NMDA) glutamate receptor mediated synaptic currents, was increased after males sang to females, and also after they saw females without singing, but not after they sang while alone. Potentiation after female exposure alone did not appear to result from stress, as it was not blocked by inhibition of glucocorticoid receptors. This potentiation was restricted to synapses of dopaminergic projection neurons, and appeared to be expressed postsynaptically. This study supports a model in which VTA dopaminergic neurons are more strongly activated during singing used for courtship than during non-courtship singing, and thus can provide social context-dependent modulation to forebrain areas. More generally, these results demonstrate that an intense social encounter can trigger the same pathways of neuronal plasticity as addictive drugs.
Conflict of interest statement
Figures
References
-
- Ikemoto S, Panksepp J. The role of nucleus accumbens dopamine in motivated behavior: a unifying interpretation with special reference to reward-seeking. Brain Res Brain Res Rev. 1999;31:6–41. - PubMed
-
- Schultz W. Behavioral theories and the neurophysiology of reward. Annu Rev Psychol. 2006;57:87–115. - PubMed
-
- Fields HL, Hjelmstad GO, Margolis EB, Nicola SM. Ventral tegmental area neurons in learned appetitive behavior and positive reinforcement. Annu Rev Neurosci. 2007;30:289–316. - PubMed
-
- Wise RA. Dopamine, learning and motivation. Nat Rev Neurosc. 2004;5:483–94. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
