

Mechanisms of nuclear vitamin D receptor resistance in Harvey-ras-transfected cells
- PMID: 18829283
- PMCID: PMC2757168
- DOI: 10.1016/j.jnutbio.2008.06.008
Mechanisms of nuclear vitamin D receptor resistance in Harvey-ras-transfected cells
Abstract
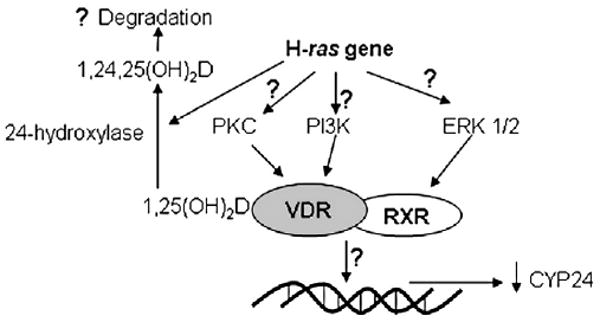
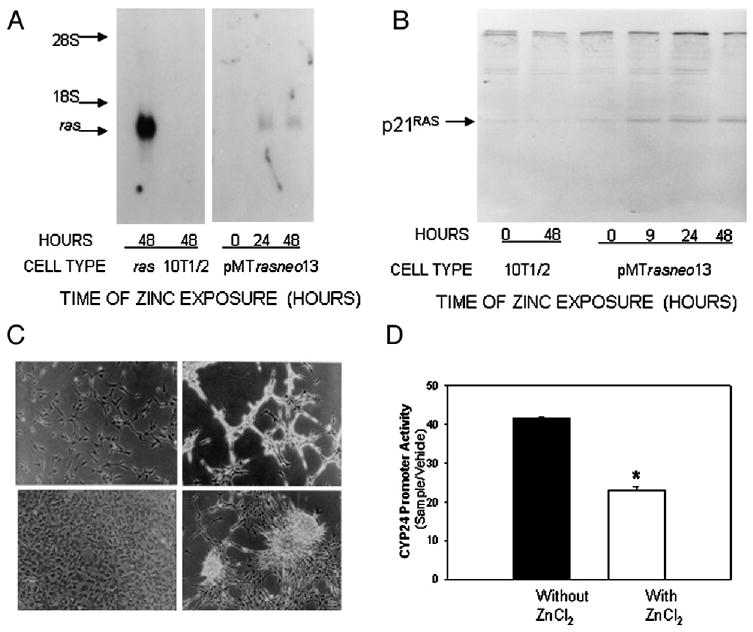
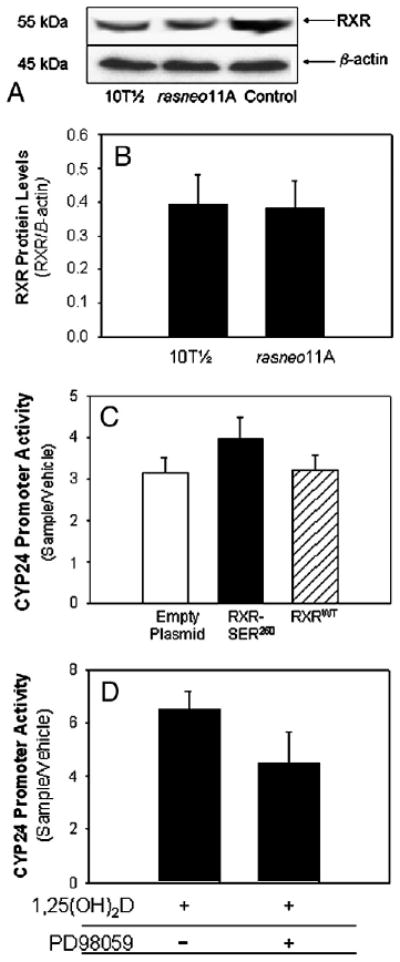
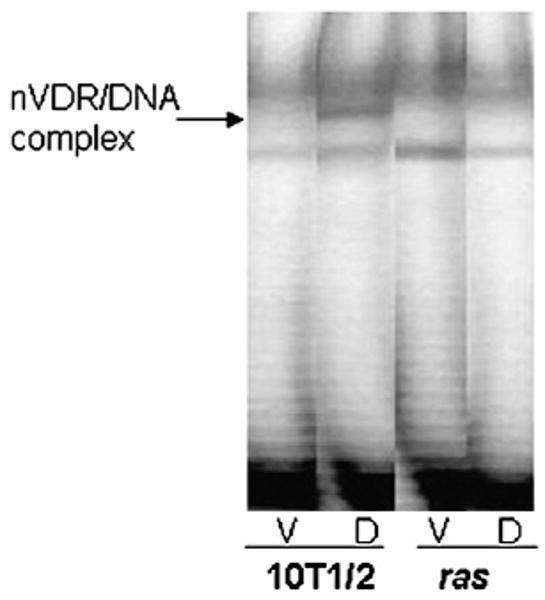
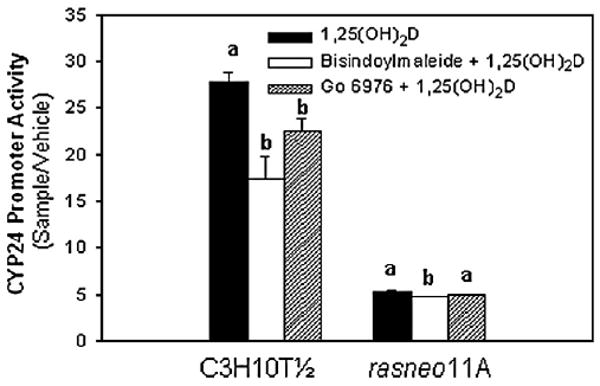
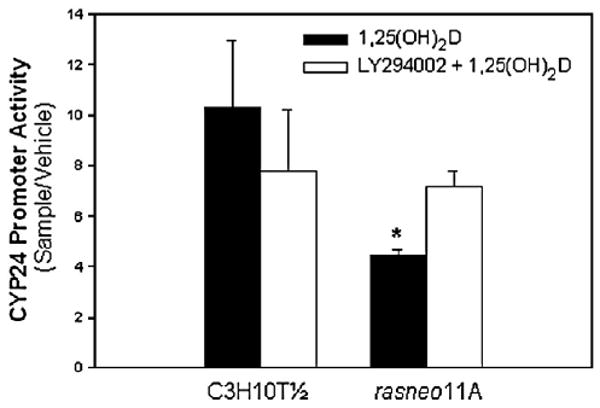
The hormone 1,25 dihydroxyvitamin D (1,25(OH)(2)D) binds to the nuclear vitamin D receptor (nVDR), which heterodimerizes with retinoid X receptor alpha (RXRalpha), and this complex interacts with specific response elements [vitamin D response elements (VDREs)] to regulate gene transcription. Previous results show a significant reduction in 1,25(OH)(2)D-induced nVDR transcriptional activity in fibroblast (C3H10T1/2) cells transfected with the Harvey ras gene (ras cells) compared with parental cells. The purpose of this study was to investigate the mechanisms by which the H-ras gene interferes with nVDR transcriptional activity. Similar to the ras cells, transcriptional activity of the nVDR was reduced following induction of the H-ras gene for 9 days. The ras cells expressed similar protein levels of RXRalpha with the parent cells, and overexpression of the wild-type RXRalpha plasmid did not restore 1,25(OH)(2)D-mediated nVDR activity in ras cells. Inhibiting activation of extracellular signal-regulated kinase (ERK1/2) had no effect on nVDR activity in ras cells. Furthermore, the binding of nVDR to VDREs was reduced in 1,25(OH)(2)D-treated ras cells. In addition, neither treatment of ras cells with an inhibitor (ketoconazole) of the 1,25(OH)(2)D degradative enzyme, 24-hydroxylase, nor the protein kinase C inhibitors, bisindoylmaleimide I and Gö 6976, had an effect on nVDR activity. In contrast, inhibition of phosphatidylinositol 3-kinase (PI3K) with LY294002 resulted in a 1.6-fold significant increase in the nVDR activity in the ras cells. Taken together, these results indicate that PI3K may, at least in part, mediate the suppression of the 1,25(OH)(2)D regulation of nVDR transcriptional activity by the H-ras gene, leading to reduced ability to associate with response elements.
Figures
Similar articles
-
Effects of MAPK signaling on 1,25-dihydroxyvitamin D-mediated CYP24 gene expression in the enterocyte-like cell line, Caco-2.J Cell Physiol. 2009 Apr;219(1):132-42. doi: 10.1002/jcp.21657. J Cell Physiol. 2009. PMID: 19097033 Free PMC article.
-
1Alpha,25-dihydroxyvitamin D3-mediated stimulation of steroid sulphatase activity in myeloid leukaemic cell lines requires VDRnuc-mediated activation of the RAS/RAF/ERK-MAP kinase signalling pathway.J Cell Biochem. 2006 Jun 1;98(3):590-617. doi: 10.1002/jcb.20787. J Cell Biochem. 2006. PMID: 16440327
-
Membrane actions of vitamin D metabolites 1alpha,25(OH)2D3 and 24R,25(OH)2D3 are retained in growth plate cartilage cells from vitamin D receptor knockout mice.J Cell Biochem. 2003 Dec 15;90(6):1207-23. doi: 10.1002/jcb.10716. J Cell Biochem. 2003. PMID: 14635194
-
New understanding of the molecular mechanism of receptor-mediated genomic actions of the vitamin D hormone.Bone. 1995 Aug;17(2 Suppl):33S-38S. doi: 10.1016/8756-3282(95)00205-r. Bone. 1995. PMID: 8579895 Review.
-
Regulation of 25-hydroxyvitamin D3-24-hydroxylase mRNA by 1,25-dihydroxyvitamin D3 and parathyroid hormone.J Cell Biochem. 2003 Feb 1;88(2):234-7. doi: 10.1002/jcb.10341. J Cell Biochem. 2003. PMID: 12520520 Review.
Cited by
-
Mechanisms of oncogene-induced genomic instability.Biophys Chem. 2017 Jun;225:49-57. doi: 10.1016/j.bpc.2016.11.008. Epub 2016 Nov 24. Biophys Chem. 2017. PMID: 28073589 Free PMC article. Review.
-
Vitamin D and Breast Cancer: Mechanistic Update.JBMR Plus. 2021 Dec 10;5(12):e10582. doi: 10.1002/jbm4.10582. eCollection 2021 Dec. JBMR Plus. 2021. PMID: 34950835 Free PMC article.
-
Apc(MIN) modulation of vitamin D secosteroid growth control.Carcinogenesis. 2010 Aug;31(8):1434-41. doi: 10.1093/carcin/bgq098. Epub 2010 May 20. Carcinogenesis. 2010. PMID: 20488884 Free PMC article.
-
1α, 25-Dihydroxyvitamin D regulates hypoxia-inducible factor-1α in untransformed and Harvey-ras transfected breast epithelial cells.Cancer Lett. 2010 Dec 8;298(2):159-66. doi: 10.1016/j.canlet.2010.06.014. Epub 2010 Jul 23. Cancer Lett. 2010. PMID: 20655141 Free PMC article.
-
1,25-dihydroxyvitamin D regulation of glucose metabolism in Harvey-ras transformed MCF10A human breast epithelial cells.J Steroid Biochem Mol Biol. 2013 Nov;138:81-9. doi: 10.1016/j.jsbmb.2013.03.012. Epub 2013 Apr 22. J Steroid Biochem Mol Biol. 2013. PMID: 23619337 Free PMC article.
References
-
- Studzinski GP, Moore DC. Sunlight — can it prevent as well as cause cancer? Cancer Res. 1995;55:4014–22. - PubMed
-
- Weaver CM, Fleet JC. Vitamin D requirements: current and future. Am J Clin Nutr. 2004;80:1735S–9S. - PubMed
-
- Narvaez CJ, Welsh J. Role of mitochondria and caspases in vitamin D-mediated apoptosis of MCF-7 breast cancer cells. J Biol Chem. 2001;276:9101–7. - PubMed
-
- Wang X, Studzinski GP. Activation of extracellular signal-regulated kinases (ERKs) defines the first phase of 1,25-dihydroxyvitamin D3-induced differentiation of HL60 cells. J Cell Biochem. 2001;80:471–82. - PubMed
-
- Lin R, Amizuka N, Sasaki T, Aarts MM, Ozawa H, Goltzman D, et al. 1Alpha,25-dihydroxyvitamin D3 promotes vascularization of the chondro-osseous junction by stimulating expression of vascular endothelial growth factor and matrix metalloproteinase 9. J Bone Miner Res. 2002;17:1604–12. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
