

Glycosylation of gp41 of simian immunodeficiency virus shields epitopes that can be targets for neutralizing antibodies
- PMID: 18829751
- PMCID: PMC2593353
- DOI: 10.1128/JVI.01382-08
Glycosylation of gp41 of simian immunodeficiency virus shields epitopes that can be targets for neutralizing antibodies
Abstract
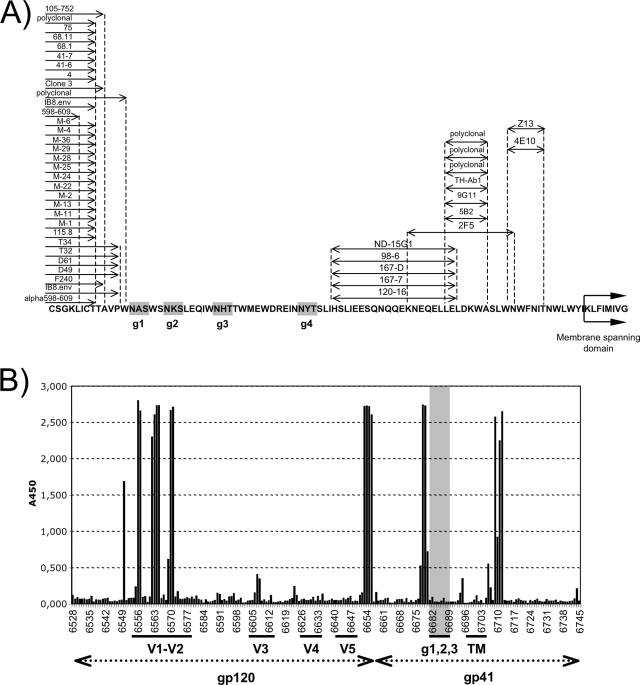
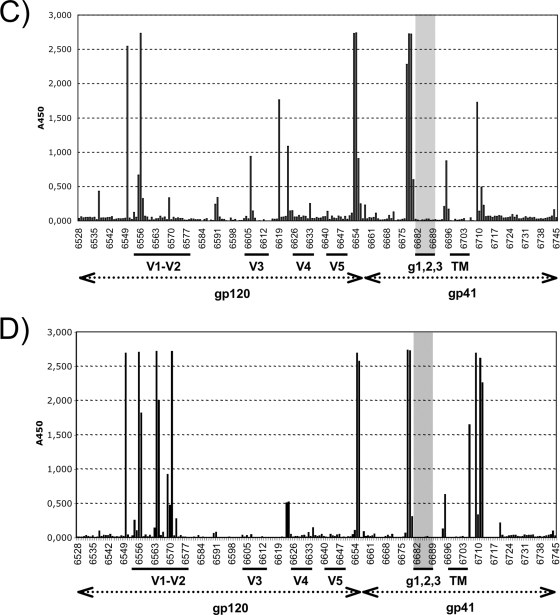
Human immunodeficiency virus type 1 and simian immunodeficiency virus possess three closely spaced, highly conserved sites for N-linked carbohydrate attachment in the extracellular domain of the transmembrane protein gp41. We infected rhesus monkeys with a variant of cloned SIVmac239 lacking the second and third sites or with a variant strain lacking all three of SIVmac239's glycosylation sites in gp41. For each mutation, asparagine (N) in the canonical N-X-S/T recognition sequence for carbohydrate attachment was changed to the structurally similar glutamine such that two nucleotide changes would be required for a reversion of the mutated codon. By 16 weeks, experimentally infected monkeys made antibodies that neutralized the mutant viruses to high titers. Such antibodies were not observed in monkeys infected with the parental virus. Thus, new specificities were revealed as a result of the carbohydrate attachment mutations, and antibodies of these specificities had neutralizing activity. Unlike monkeys infected with the parental virus, monkeys infected with the mutant viruses made antibodies that reacted with peptides corresponding to the sequences in this region. Furthermore, there was strong selective pressure for the emergence of variant sequences in this region during the course of infection. By analyzing the neutralization profiles of sequence variants, we were able to define three mutations (Q625R, K631N, and Q634H) in the region of the glycosylation site mutations that conferred resistance to neutralization by plasma from the monkeys infected with mutant virus. Based on the reactivity of antibodies to peptides in this region and the colocalization of neutralization escape mutations, we conclude that N-linked carbohydrates in the ectodomain of the transmembrane protein shield underlying epitopes that would otherwise be the direct targets of neutralizing antibodies.
Figures
References
-
- Back, N. K. T., L. Smit, J.-J. De Jong, W. Keulen, M. Schutten, J. Goudsmit, and M. Tersmette. 1994. An N-glycan within the human immunodeficiency virus type 1 gp120 V3 loop affects virus neutralization. Virology 199431-438. - PubMed
-
- Bahraoui, E., A. Benjouad, D. Guetard, H. Kolbe, J. C. Gluckman, and L. Montagnier. 1992. Study of the interaction of HIV-1 and HIV-2 envelope glycoproteins with the CD4 receptor and role of N-glycans. AIDS Res. Hum. Retrovir. 8565-573. - PubMed
-
- Binley, J. M., R. Wyatt, E. Desjardins, P. D. Kwong, W. Hendrickson, J. P. Moore, and J. Sodroski. 1998. Analysis of the interaction of antibodies with a conserved enzymatically deglycosylated core of the HIV type 1 envelope glycoprotein 120. AIDS Res. Hum. Retrovir. 14191-198. - PubMed
-
- Blay, W. M., S. Gnanakaran, B. Foley, N. A. Doria-Rose, B. T. Korber, and N. L. Haigwood. 2006. Consistent patterns of change during the divergence of human immunodeficiency virus type 1 envelope from that of the inoculated virus in simian/human immunodeficiency virus-infected macaques. J. Virol. 80999-1014. - PMC - PubMed
-
- Bolmstedt, A., S. Sjolander, J. E. Hansen, L. Akerblom, A. Hemming, S. L. Hu, B. Morein, and S. Olofsson. 1996. Influence of N-linked glycans in V4-V5 region of human immunodeficiency virus type 1 glycoprotein gp160 on induction of a virus-neutralizing humoral response. J. Acquir. Immune Defic. Syndr. Hum. Retrovirol. 12213-220. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
