

S-adenosyl homocysteine-induced hyperpolyadenylation of vesicular stomatitis virus mRNA requires the methyltransferase activity of L protein
- PMID: 18829753
- PMCID: PMC2593356
- DOI: 10.1128/JVI.01225-08
S-adenosyl homocysteine-induced hyperpolyadenylation of vesicular stomatitis virus mRNA requires the methyltransferase activity of L protein
Abstract
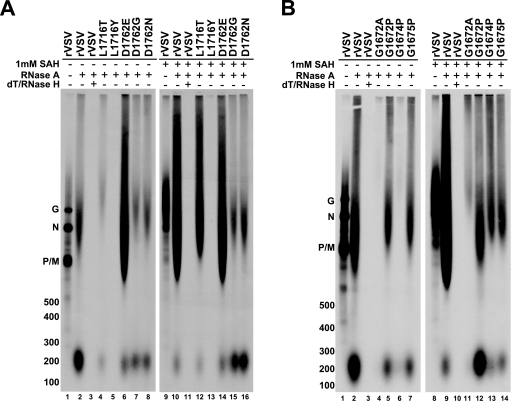
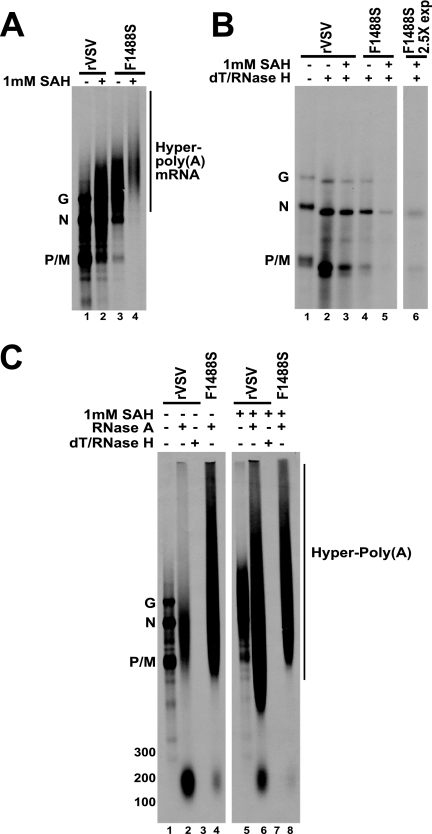
There are many unique aspects of vesicular stomatitis virus (VSV) transcription. In addition to its unusual mRNA capping and methyltransferase mechanisms, the addition of S-adenosyl homocysteine (SAH), which is the by-product and competitive inhibitor of S-adenosyl methionine (SAM)-mediated methyltransferase reactions, leads to synthesis of poly(A) tails on the 3' end of VSV mRNAs that are 10- or 20-fold longer than normal. The mechanism by which this occurs is not understood, since it has been shown that productive transcription is not dependent on 5' cap methylation and full-length VSV mRNAs can be synthesized in the absence of SAM. To investigate this unusual phenotype, we assayed the effects of SAH on transcription using a panel of recombinant viruses that contained mutations in domain VI of the VSV L protein. The L proteins we investigated displayed a range of 5' cap methyltransferase activities. In the present study, we show that the ability of the VSV L protein to catalyze methyl transfer correlates with its sensitivity to SAH with respect to polyadenylation, thereby indicating an intriguing connection between 5' and 3' end mRNA modifications. We also identified an L protein mutant that hyperpolyadenylates mRNA irrespective of the presence or absence of exogenous SAH. Further, the data presented here show that the wild-type L protein hyperpolyadenylates a percentage of VSV mRNAs in infected cells as well as in vitro.
Figures
Similar articles
-
Oligomerization of the Vesicular Stomatitis Virus Phosphoprotein Is Dispensable for mRNA Synthesis but Facilitates RNA Replication.J Virol. 2020 Jun 16;94(13):e00115-20. doi: 10.1128/JVI.00115-20. Print 2020 Jun 16. J Virol. 2020. PMID: 32321813 Free PMC article.
-
A freeze frame view of vesicular stomatitis virus transcription defines a minimal length of RNA for 5' processing.PLoS Pathog. 2011 Jun;7(6):e1002073. doi: 10.1371/journal.ppat.1002073. Epub 2011 Jun 2. PLoS Pathog. 2011. PMID: 21655110 Free PMC article.
-
Ribose 2'-O methylation of the vesicular stomatitis virus mRNA cap precedes and facilitates subsequent guanine-N-7 methylation by the large polymerase protein.J Virol. 2009 Nov;83(21):11043-50. doi: 10.1128/JVI.01426-09. Epub 2009 Aug 26. J Virol. 2009. PMID: 19710136 Free PMC article.
-
An unconventional pathway of mRNA cap formation by vesiculoviruses.Virus Res. 2011 Dec;162(1-2):100-9. doi: 10.1016/j.virusres.2011.09.012. Epub 2011 Sep 16. Virus Res. 2011. PMID: 21945214 Free PMC article. Review.
-
[The multifunctional RNA polymerase L protein of non-segmented negative strand RNA viruses catalyzes unique mRNA capping].Uirusu. 2014;64(2):165-78. doi: 10.2222/jsv.64.165. Uirusu. 2014. PMID: 26437839 Review. Japanese.
Cited by
-
Opposing effects of inhibiting cap addition and cap methylation on polyadenylation during vesicular stomatitis virus mRNA synthesis.J Virol. 2009 Feb;83(4):1930-40. doi: 10.1128/JVI.02162-08. Epub 2008 Dec 10. J Virol. 2009. PMID: 19073725 Free PMC article.
-
Transcriptional Control and mRNA Capping by the GDP Polyribonucleotidyltransferase Domain of the Rabies Virus Large Protein.Viruses. 2019 Jun 1;11(6):504. doi: 10.3390/v11060504. Viruses. 2019. PMID: 31159413 Free PMC article. Review.
-
Components and Architecture of the Rhabdovirus Ribonucleoprotein Complex.Viruses. 2020 Aug 29;12(9):959. doi: 10.3390/v12090959. Viruses. 2020. PMID: 32872471 Free PMC article. Review.
-
RNA Synthesis and Capping by Non-segmented Negative Strand RNA Viral Polymerases: Lessons From a Prototypic Virus.Front Microbiol. 2019 Jul 10;10:1490. doi: 10.3389/fmicb.2019.01490. eCollection 2019. Front Microbiol. 2019. PMID: 31354644 Free PMC article. Review.
-
Identification of aromatic amino acid residues in conserved region VI of the large polymerase of vesicular stomatitis virus is essential for both guanine-N-7 and ribose 2'-O methyltransferases.Virology. 2010 Dec 20;408(2):241-52. doi: 10.1016/j.virol.2010.09.017. Epub 2010 Oct 18. Virology. 2010. PMID: 20961592 Free PMC article.
References
-
- Barr, J. N., X. Tang, E. Hinzman, R. Shen, and G. W. Wertz. 2008. The VSV polymerase can initiate at mRNA start sites located either up or downstream of a transcription termination signal but size of the intervening intergenic region affects efficiency of initiation. Virology 374361-370. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
