

The scaffold protein POSH regulates axon outgrowth
- PMID: 18829867
- PMCID: PMC2592661
- DOI: 10.1091/mbc.e08-02-0231
The scaffold protein POSH regulates axon outgrowth
Abstract
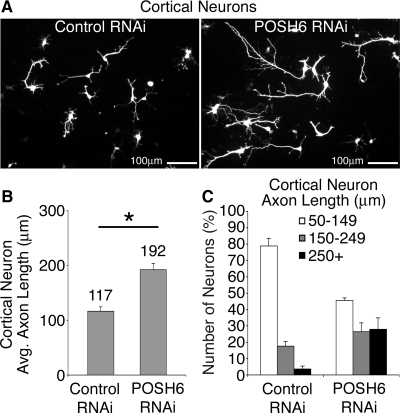
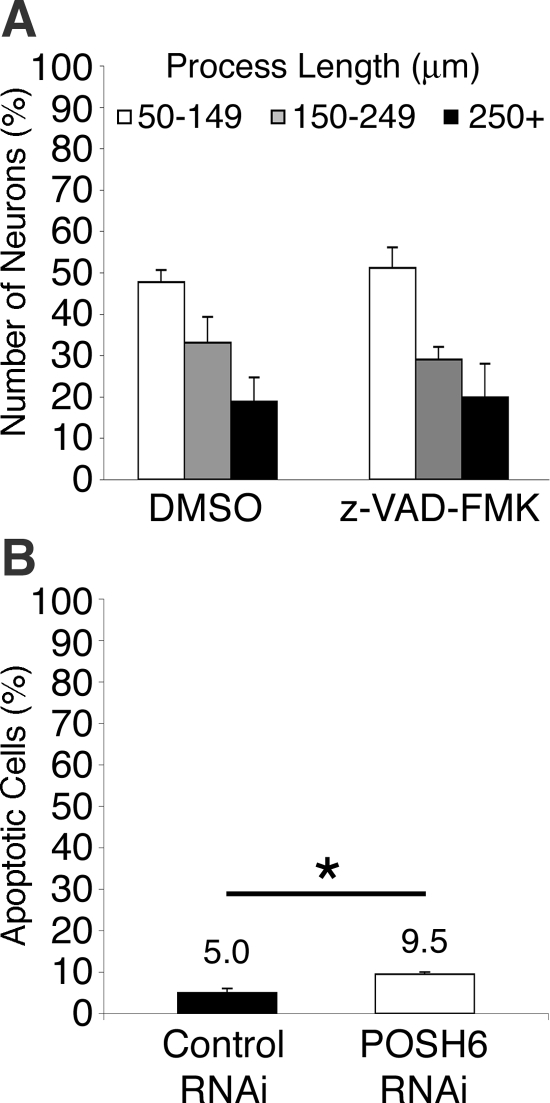
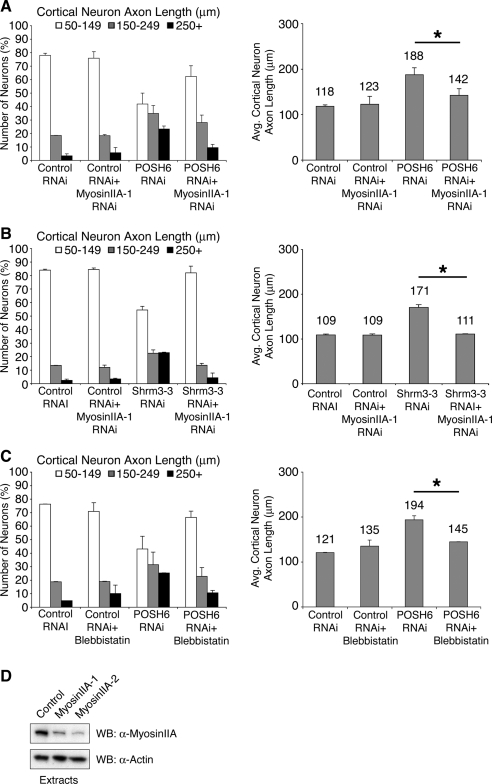
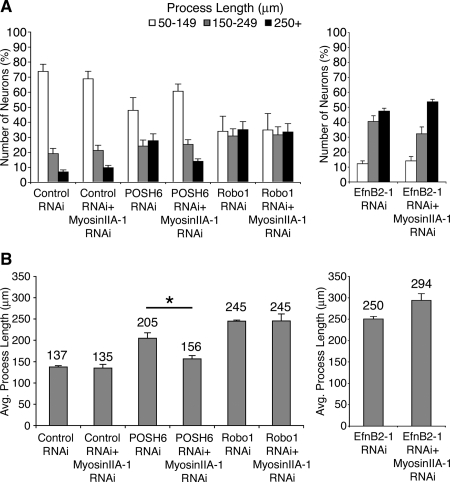
How scaffold proteins integrate signaling pathways with cytoskeletal components to drive axon outgrowth is not well understood. We report here that the multidomain scaffold protein Plenty of SH3s (POSH) regulates axon outgrowth. Reduction of POSH function by RNA interference (RNAi) enhances axon outgrowth in differentiating mouse primary cortical neurons and in neurons derived from mouse P19 cells, suggesting POSH negatively regulates axon outgrowth. Complementation analysis reveals a requirement for the third Src homology (SH) 3 domain of POSH, and we find that the actomyosin regulatory protein Shroom3 interacts with this domain of POSH. Inhibition of Shroom3 expression by RNAi leads to increased process lengths, as observed for POSH RNAi, suggesting that POSH and Shroom function together to inhibit process outgrowth. Complementation analysis and interference of protein function by dominant-negative approaches suggest that Shroom3 recruits Rho kinase to inhibit process outgrowth. Furthermore, inhibition of myosin II function reverses the POSH or Shroom3 RNAi phenotype, indicating a role for myosin II regulation as a target of the POSH-Shroom complex. Collectively, these results suggest that the molecular scaffold protein POSH assembles an inhibitory complex that links to the actin-myosin network to regulate neuronal process outgrowth.
Figures






References
-
- Brown J., Bridgman P. C. Role of myosin II in axon outgrowth. J. Histochem. Cytochem. 2003;51:421–428. - PubMed
-
- Chantler P. D., Wylie S. R. Elucidation of the separate roles of myosins IIA and IIB during neurite outgrowth, adhesion and retraction. IEE Proc. Nanobiotechnol. 2003;150:111–125. - PubMed
-
- Dietz M. L., Bernaciak T. M., Vendetti F., Kielec J. M., Hildebrand J. D. Differential actin-dependent localization modulates the evolutionarily conserved activity of Shroom family proteins. J. Biol. Chem. 2006;281:20542–20554. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
