

Decreased expression of cytosolic pyruvate kinase in potato tubers leads to a decline in pyruvate resulting in an in vivo repression of the alternative oxidase
- PMID: 18829984
- PMCID: PMC2577264
- DOI: 10.1104/pp.108.126516
Decreased expression of cytosolic pyruvate kinase in potato tubers leads to a decline in pyruvate resulting in an in vivo repression of the alternative oxidase
Abstract
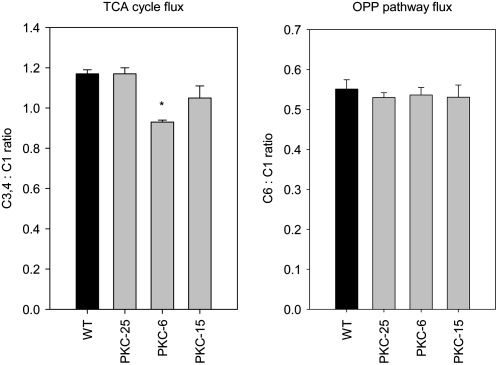
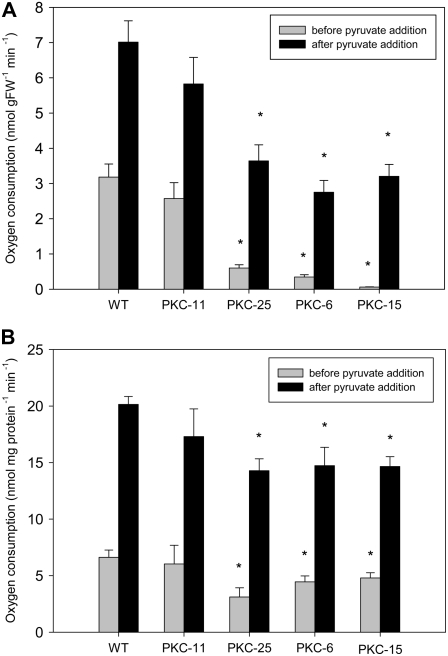
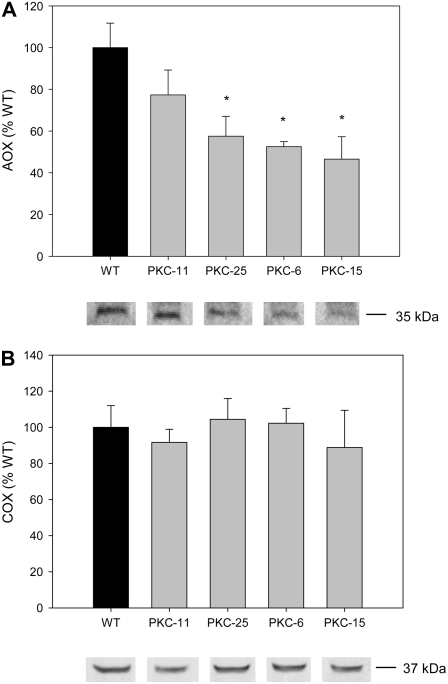
The aim of this work was to investigate the effect of decreased cytosolic pyruvate kinase (PKc) on potato (Solanum tuberosum) tuber metabolism. Transgenic potato plants with strongly reduced levels of PKc were generated by RNA interference gene silencing under the control of a tuber-specific promoter. Metabolite profiling showed that decreased PKc activity led to a decrease in the levels of pyruvate and some other organic acids involved in the tricarboxylic acid cycle. Flux analysis showed that this was accompanied by changes in carbon partitioning, with carbon flux being diverted from glycolysis toward starch synthesis. However, this metabolic shift was relatively small and hence did not result in enhanced starch levels in the tubers. Although total respiration rates and the ATP to ADP ratio were largely unchanged, transgenic tubers showed a strong decrease in the levels of alternative oxidase (AOX) protein and a corresponding decrease in the capacity of the alternative pathway of respiration. External feeding of pyruvate to tuber tissue or isolated mitochondria resulted in activation of the AOX pathway, both in the wild type and the PKc transgenic lines, providing direct evidence for the regulation of AOX by changes in pyruvate levels. Overall, these results provide evidence for a crucial role of PKc in the regulation of pyruvate levels as well as the level of the AOX in heterotrophic plant tissue, and furthermore reveal that these parameters are interlinked in vivo.
Figures
Similar articles
-
Metabolic and developmental adaptations of growing potato tubers in response to specific manipulations of the adenylate energy status.Plant Physiol. 2008 Apr;146(4):1579-98. doi: 10.1104/pp.108.115758. Epub 2008 Feb 27. Plant Physiol. 2008. PMID: 18305207 Free PMC article.
-
Tuber-specific cytosolic expression of a bacterial phosphoglucomutase in potato (Solanum tuberosum L.) dramatically alters carbon partitioning.Plant Cell Physiol. 2005 Apr;46(4):588-97. doi: 10.1093/pcp/pci058. Epub 2005 Feb 2. Plant Cell Physiol. 2005. PMID: 15695443
-
Evidence of the crucial role of sucrose synthase for sink strength using transgenic potato plants (Solanum tuberosum L.).Plant J. 1995 Jan;7(1):97-107. doi: 10.1046/j.1365-313x.1995.07010097.x. Plant J. 1995. PMID: 7894514
-
Alteration of the interconversion of pyruvate and malate in the plastid or cytosol of ripening tomato fruit invokes diverse consequences on sugar but similar effects on cellular organic acid, metabolism, and transitory starch accumulation.Plant Physiol. 2013 Feb;161(2):628-43. doi: 10.1104/pp.112.211094. Epub 2012 Dec 18. Plant Physiol. 2013. PMID: 23250627 Free PMC article.
-
Transgenic 14-3-3 isoforms in plants: the metabolite profiling of repressed 14-3-3 protein synthesis in transgenic potato plants.Biochem Soc Trans. 2002 Aug;30(4):405-10. doi: 10.1042/bst0300405. Biochem Soc Trans. 2002. PMID: 12196104 Review.
Cited by
-
Isolation and Characterization of DkPK Genes Associated with Natural Deastringency in C-PCNA Persimmon.Front Plant Sci. 2016 Feb 17;7:156. doi: 10.3389/fpls.2016.00156. eCollection 2016. Front Plant Sci. 2016. PMID: 26925075 Free PMC article.
-
Identification and computational annotation of genes differentially expressed in pulp development of Cocos nucifera L. by suppression subtractive hybridization.BMC Plant Biol. 2014 Aug 2;14:205. doi: 10.1186/s12870-014-0205-7. BMC Plant Biol. 2014. PMID: 25084812 Free PMC article.
-
DkPK Genes Promote Natural Deastringency in C-PCNA Persimmon by Up-regulating DkPDC and DkADH Expression.Front Plant Sci. 2017 Feb 13;8:149. doi: 10.3389/fpls.2017.00149. eCollection 2017. Front Plant Sci. 2017. PMID: 28243247 Free PMC article.
-
A metabolomic approach to study major metabolite changes during acclimation to limiting CO2 in Chlamydomonas reinhardtii.Plant Physiol. 2010 Sep;154(1):187-96. doi: 10.1104/pp.110.157651. Epub 2010 Jul 15. Plant Physiol. 2010. PMID: 20634393 Free PMC article.
-
Integrative response of plant mitochondrial electron transport chain to nitrogen source.Plant Cell Rep. 2011 Feb;30(2):195-204. doi: 10.1007/s00299-010-0955-0. Epub 2010 Dec 4. Plant Cell Rep. 2011. PMID: 21132432 Review.
References
-
- Arabidopsis Genome Initiative (2000) Analysis of the genome sequence of the flowering plant Arabidopsis thaliana. Nature 408 796–815 - PubMed
-
- Baud S, Wuilleme S, Dubreucq B, de Almeida A, Vuagnat C, Lepiniec L, Miquel M, Rochat C (2007) Function of plastidial pyruvate kinases in seeds of Arabidopsis thaliana. Plant J 52 405–419 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
