

Structural analysis of fish versus mammalian hemoglobins: effect of the heme pocket environment on autooxidation and hemin loss
- PMID: 18831041
- PMCID: PMC2649966
- DOI: 10.1002/prot.22236
Structural analysis of fish versus mammalian hemoglobins: effect of the heme pocket environment on autooxidation and hemin loss
Abstract
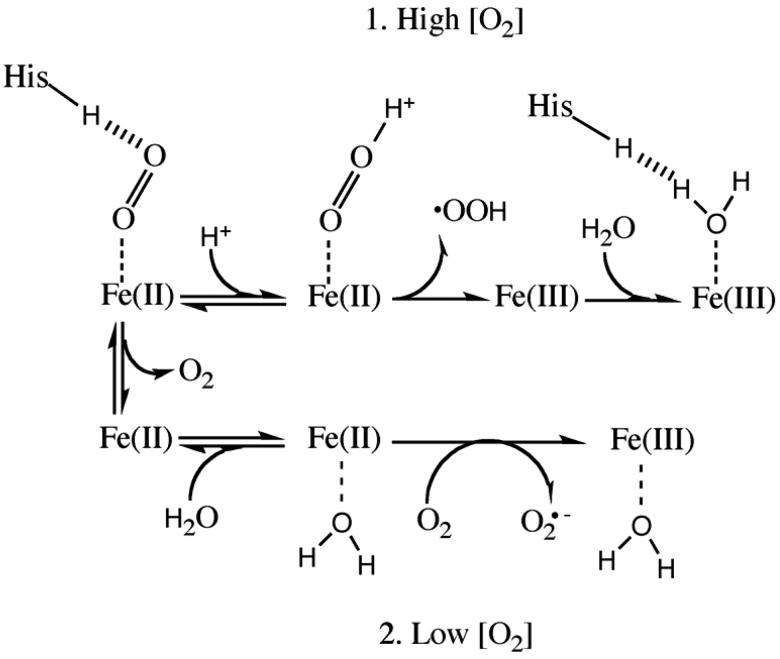
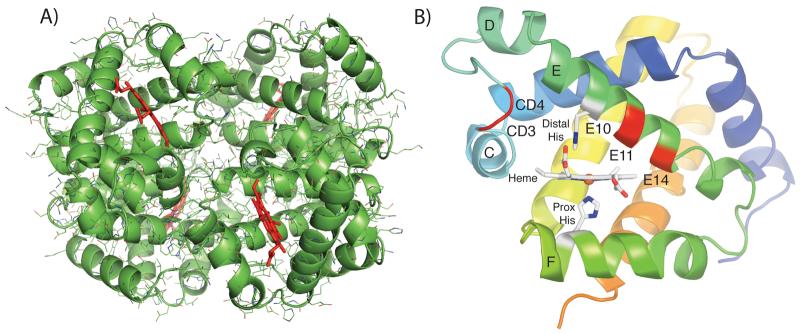
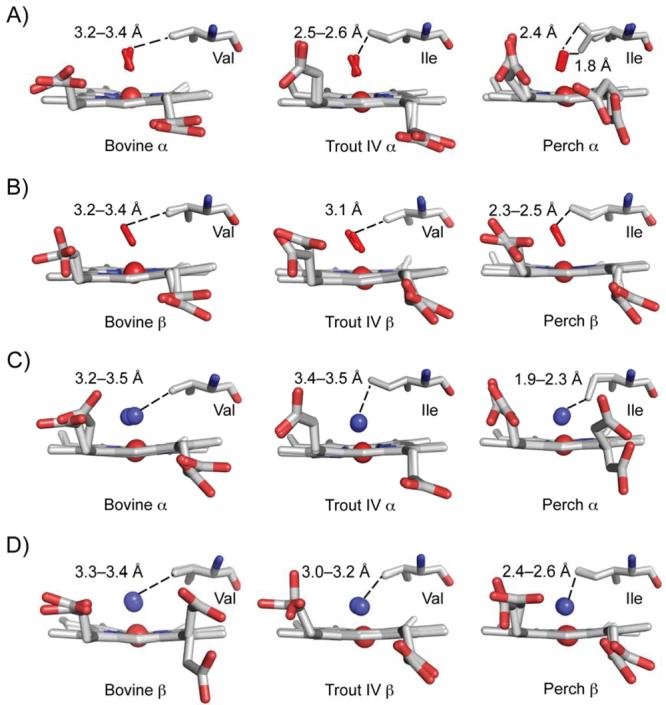
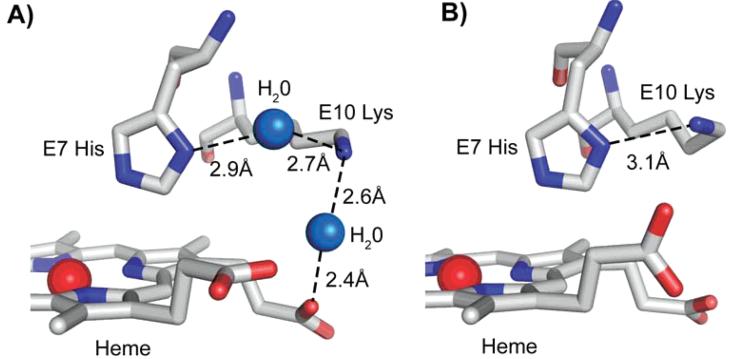
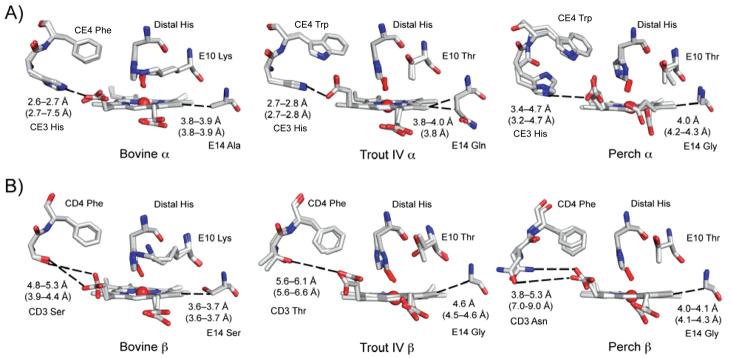
The underlying stereochemical mechanisms for the dramatic differences in autooxidation and hemin loss rates of fish versus mammalian hemoglobins (Hb) have been examined by determining the crystal structures of perch, trout IV, and bovine Hb at high and low pH. The fish Hbs autooxidize and release hemin approximately 50- to 100-fold more rapidly than bovine Hb. Five specific amino acid replacements in the CD corner and along the E helix appear to cause the increased susceptibility of fish Hbs to oxidative degradation compared with mammalian Hbs. Ile is present at the E11 helical position in most fish Hb chains whereas a smaller Val residue is present in all mammalian alpha and beta chains. The larger IleE11 side chain sterically hinders bound O(2) and facilitates dissociation of the neutral superoxide radical, enhancing autooxidation. Lys(E10) is found in most mammalian Hb and forms favorable electrostatic and hydrogen bonding interactions with the heme-7-propionate. In contrast, Thr(E10) is present in most fish Hbs and is too short to stabilize bound heme, and causes increased rates of hemin dissociation. Especially high rates of hemin loss in perch Hb are also due to a lack of electrostatic interaction between His(CE3) and the heme-6 propionate in alpha subunits whereas this interaction does occur in trout IV and bovine Hb. There is also a larger gap for solvent entry into the heme crevice near beta CD3 in the perch Hb (approximately 8 A) compared with trout IV Hb (approximately 6 A) which in turn is significantly higher than that in bovine Hb (approximately 4 A) at low pH. The amino acids at CD4 and E14 differ between bovine and the fish Hbs and have the potential to modulate oxidative degradation by altering the orientation of the distal histidine and the stability of the E-helix. Generally rapid rates of lipid oxidation in fish muscle can be partly attributed to the fact that fish Hbs are highly susceptible to oxidative degradation.
(c) 2008 Wiley-Liss, Inc.
Figures
References
-
- Alayash AI, Brockner Ryan BA, Eich RF, Olson JS, Cashon RE. Reactions of sperm whale myoglobin with hydrogen peroxide. Effects of distal pocket mutations on the formation and stability of the ferryl intermediate. J Biol Chem. 1999;274:2029–2037. - PubMed
-
- Trent RJ, Webster B, Bowden DK, Gilbert A, Holl PJ, Lindeman R, Lammi A, Rowell J, Hinchcliffe M, Colley A, Wilson M, Saleh M, Blackwell J, Petrou V. Complex phenotypes in the haemoglobinopathies: recommendations on screening and DNA testing. Pathology. 2006;38(6):507–519. - PubMed
-
- Umbreit J. Methemoglobin--it's not just blue: a concise review. Am J Hematol. 2007;82(2):134–144. - PubMed
-
- Watanabe J, Chou KJ, Liao JC, Miao Y, Meng HH, Ge H, Grijalva V, Hama S, Kozak K, Buga G, Whitelegge JP, Lee TD, Farias-Eisner R, Navab M, Fogelman AM, Reddy ST. Differential association of hemoglobin with proinflammatory high density lipoproteins in atherogenic/hyperlipidemic mice. A novel biomarker of atherosclerosis. J Biol Chem. 2007;282(32):23698–23707. - PubMed
-
- Grinshtein N, Bamm VV, Tsemakhovich VA, Shaklai N. Mechanism of low-density lipoprotein oxidation by hemoglobin-derived iron. Biochemistry. 2003;42(23):6977–6985. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
