

Gap junctions in the control of vascular function
- PMID: 18831678
- PMCID: PMC2933153
- DOI: 10.1089/ars.2008.2117
Gap junctions in the control of vascular function
Abstract
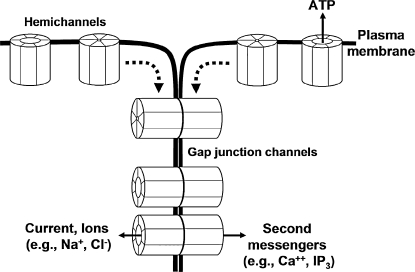
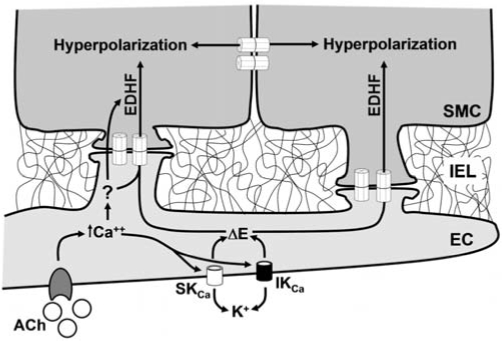
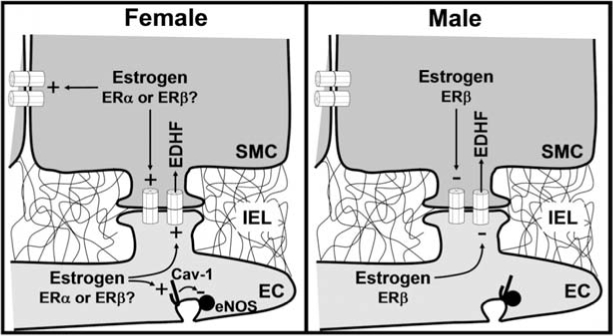
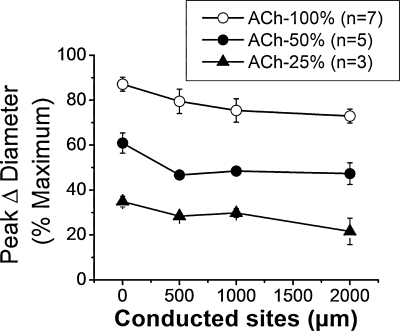
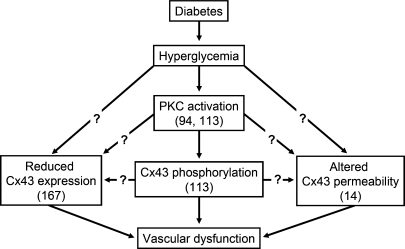
Direct intercellular communication via gap junctions is critical in the control and coordination of vascular function. In the cardiovascular system, gap junctions are made up of one or more of four connexin proteins: Cx37, Cx40, Cx43, and Cx45. The expression of more than one gap-junction protein in the vasculature is not redundant. Rather, vascular connexins work in concert, first during the development of the cardiovascular system, and then in integrating smooth muscle and endothelial cell function, and in coordinating cell function along the length of the vessel wall. In addition, connexin-based channels have emerged as an important signaling pathway in the astrocyte-mediated neurovascular coupling. Direct electrical communication between endothelial cells and vascular smooth muscle cells via gap junctions is thought to play a relevant role in the control of vasomotor tone, providing the signaling pathway known as endothelium-derived hyperpolarizing factor (EDHF). Consistent with the importance of gap junctions in the regulation of vasomotor tone and arterial blood pressure, the expression of connexins is altered in diseases associated with vascular complications. In this review, we discuss the participation of connexin-based channels in the control of vascular function in physiologic and pathologic conditions, with a special emphasis on hypertension and diabetes.
Figures
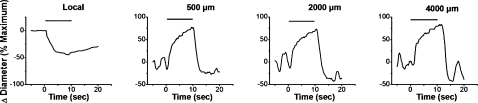
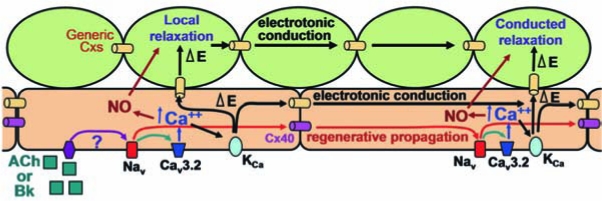
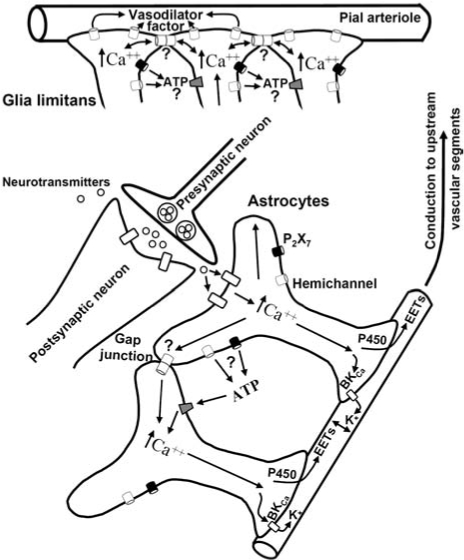
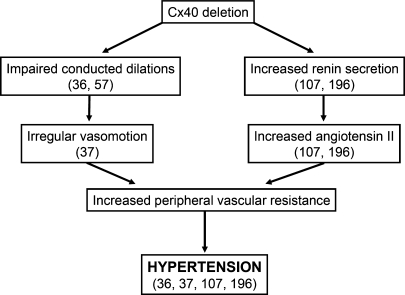
References
-
- Ahluwalia A. Hobbs AJ. Endothelium-derived C-type natriuretic peptide: more than just a hyperpolarizing factor. Trends Pharmacol Sci. 2005;26:162–167. - PubMed
-
- Anderson CM. Nedergaard M. Astrocyte-mediated control of cerebral microcirculation. Trends Neurosci. 2003;26:340–344. - PubMed
-
- Andresen J. Shafi NI. Bryan RM., Jr Endothelial influences on cerebrovascular tone. J Appl Physiol. 2006;100:318–327. - PubMed
-
- Archer SL. Gragasin FS. Wu X. Wang S. McMurtry S. Kim DH. Platonov M. Koshal A. Hashimoto K. Campbell WB. Falck JR. Michelakis ED. Endothelium-derived hyperpolarizing factor in human internal mammary artery is 11,12-epoxyeicosatrienoic acid and causes relaxation by activating smooth muscle BK(Ca) channels. Circulation. 2003;107:769–776. - PubMed
-
- Arensbak B. Mikkelsen HB. Gustafsson F. Christensen T. Holstein-Rathlou NH. Expression of connexin 37, 40, and 43 mRNA and protein in renal preglomerular arterioles. Histochem Cell Biol. 2001;115:479–487. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
