

The Mycoplasma pneumoniae MPN229 gene encodes a protein that selectively binds single-stranded DNA and stimulates Recombinase A-mediated DNA strand exchange
- PMID: 18831760
- PMCID: PMC2572620
- DOI: 10.1186/1471-2180-8-167
The Mycoplasma pneumoniae MPN229 gene encodes a protein that selectively binds single-stranded DNA and stimulates Recombinase A-mediated DNA strand exchange
Abstract
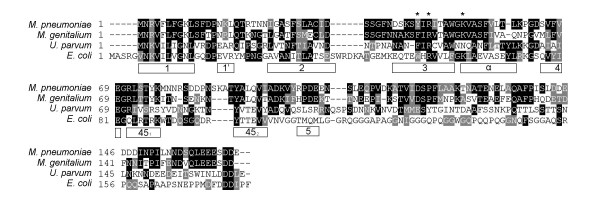
Background: Mycoplasma pneumoniae has previously been characterized as a micro-organism that is genetically highly stable. In spite of this genetic stability, homologous DNA recombination has been hypothesized to lie at the basis of antigenic variation of the major surface protein, P1, of M. pneumoniae. In order to identify the proteins that may be involved in homologous DNA recombination in M. pneumoniae, we set out to characterize the MPN229 open reading frame (ORF), which bears sequence similarity to the gene encoding the single-stranded DNA-binding (SSB) protein of other micro-organisms.
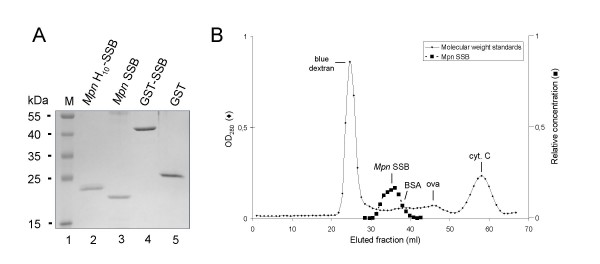
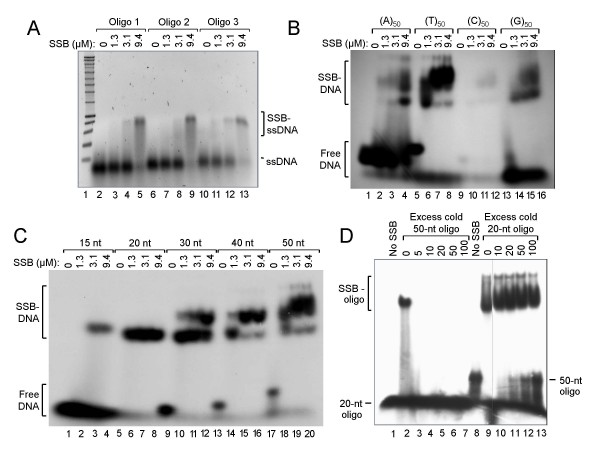
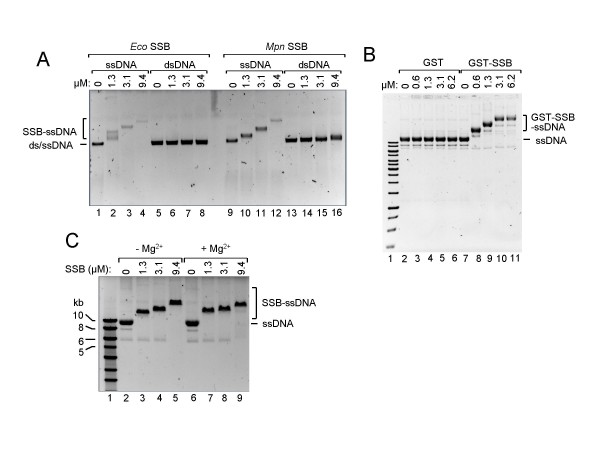
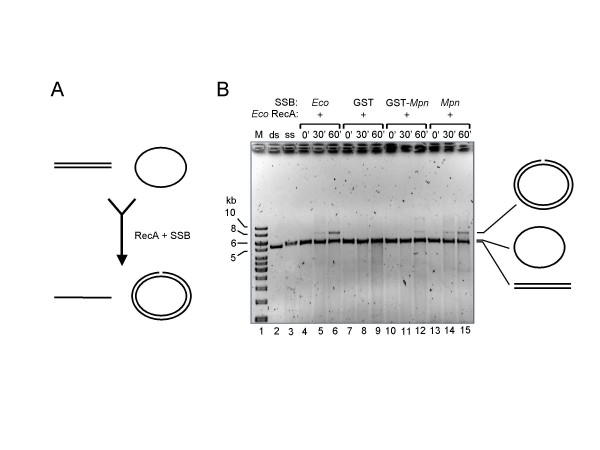
Results: The MPN229 ORF has the capacity to encode a 166-amino acid protein with a calculated molecular mass of 18.4 kDa. The amino acid sequence of this protein (Mpn SSB) is most closely related to that of the protein predicted to be encoded by the MG091 gene from Mycoplasma genitalium (61% identity). The MPN229 ORF was cloned, and different versions of Mpn SSB were expressed in E. coli and purified to > 95% homogeneity. The purified protein was found to exist primarily as a homo-tetramer in solution, and to strongly and selectively bind single-stranded DNA (ssDNA) in a divalent cation- and DNA substrate sequence-independent manner. Mpn SSB was found to bind with a higher affinity to ssDNA substrates larger than 20 nucleotides than to smaller substrates. In addition, the protein strongly stimulated E. coli Recombinase A (RecA)-promoted DNA strand exchange, which indicated that Mpn SSB may play an important role in DNA recombination processes in M. pneumoniae.
Conclusion: The M. pneumoniae MPN229 gene encodes a protein, Mpn SSB, which selectively and efficiently binds ssDNA, and stimulates E. coli RecA-promoted homologous DNA recombination. Consequently, the Mpn SSB protein may play a crucial role in DNA recombinatorial pathways in M. pneumoniae. The results from this study will pave the way for unraveling these pathways and assess their role in antigenic variation of M. pneumoniae.
Figures
References
-
- Dandekar T, Huynen M, Regula JT, Ueberle B, Zimmermann CU, Andrade MA, Doerks T, Sanchez-Pulido L, Snel B, Suyama M, et al. Re-annotating the Mycoplasma pneumoniae genome sequence: adding value, function and reading frames. Nucleic Acids Res. 2000;28:3278–3288. doi: 10.1093/nar/28.17.3278. - DOI - PMC - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
