

G-quadruplexes: the beginning and end of UTRs
- PMID: 18832370
- PMCID: PMC2577360
- DOI: 10.1093/nar/gkn511
G-quadruplexes: the beginning and end of UTRs
Abstract
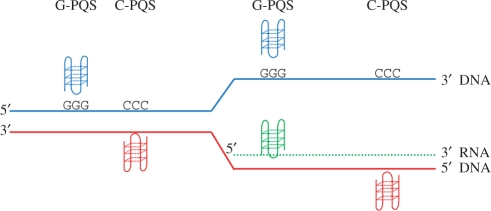
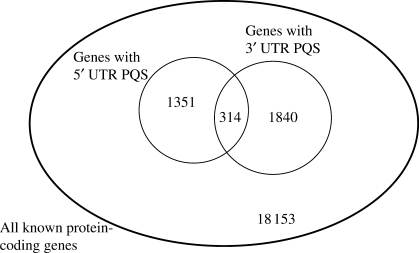
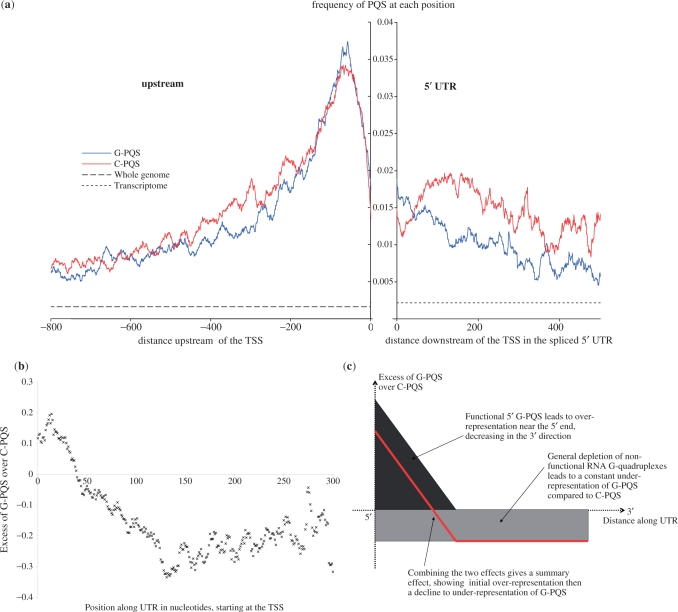
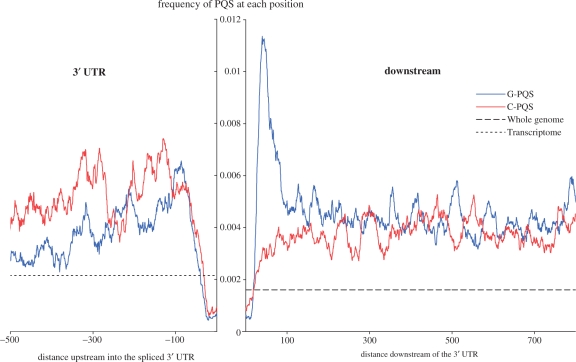
Molecular mechanisms that regulate gene expression can occur either before or after transcription. The information for post-transcriptional regulation can lie within the sequence or structure of the RNA transcript and it has been proposed that G-quadruplex nucleic acid sequence motifs may regulate translation as well as transcription. Here, we have explored the incidence of G-quadruplex motifs in and around the untranslated regions (UTRs) of mRNA. We observed a significant strand asymmetry, consistent with a general depletion of G-quadruplex-forming RNA. We also observed a positional bias in two distinct regions, each suggestive of a specific function. We observed an excess of G-quadruplex motifs towards the 5'-ends of 5'-UTRs, supportive of a hypothesis linking 5'-UTR RNA G-quadruplexes to translational control. We then analysed the vicinity of 3'-UTRs and observed an over-representation of G-quadruplex motifs immediately after the 3'-end of genes, especially in those cases where another gene is in close proximity, suggesting that G-quadruplexes may be involved in the termination of gene transcription.
Figures

References
-
- Pesole G, Mignone F, Gissi C, Grillo G, Licciulli F, Liuni S. Structural and functional features of eukaryotic mRNA untranslated regions. Gene. 2001;276:73–81. - PubMed
-
- Bartel DP. MicroRNAs: genomics, biogenesis, mechanism and function. Cell. 2004;116:281–297. - PubMed
-
- Wilkie GS, Dickson KS, Gray NK. Regulation of mRNA translation by 5′ and 3′-UTR-binding factors. Trends Bioc. Sci. 2003;28:182–188. - PubMed
-
- Mandal M, Breaker RR. Gene regulation by riboswitches. Nat. Rev. Mol. Cell Biol. 2004;5:451–463. - PubMed
-
- Kozak M. Structural features in eukaryotic mRNAs that modulate the initiation of translation. J. Biol. Chem. 1991;266:19867–19870. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
