

Dissociation of in vitro DNA deamination activity and physiological functions of AID mutants
- PMID: 18832469
- PMCID: PMC2572969
- DOI: 10.1073/pnas.0806641105
Dissociation of in vitro DNA deamination activity and physiological functions of AID mutants
Abstract
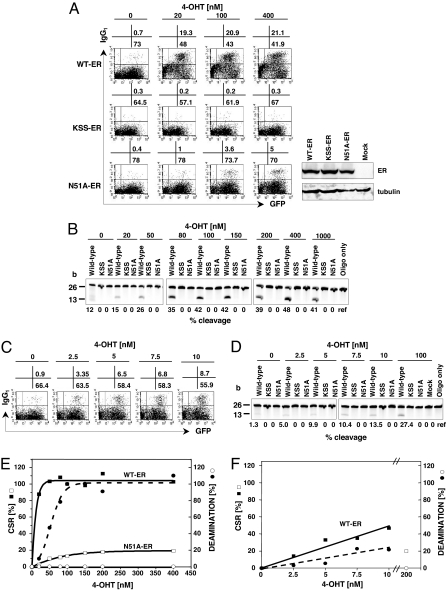
Activation-induced cytidine deaminase (AID) is essential for the DNA cleavage that initiates both somatic hypermutation (SHM) and class switch recombination (CSR) of the Ig gene. Two alternative mechanisms of DNA cleavage by AID have been proposed: RNA editing and DNA deamination. In support of the latter, AID has DNA deamination activity in cell-free systems that is assumed to represent its physiological function. To test this hypothesis, we generated various mouse AID mutants and compared their DNA deamination, CSR, and SHM activities. Here, we compared DNA deamination, CSR, and SHM activities of various AID mutants and found that most of their CSR or SHM activities were disproportionate with their DNA deamination activities. Specifically, we identified a cluster of mutants (H48A, L49A, R50A, and N51A) with low DNA deamination activity but relatively intact CSR activity. Of note is an AID mutant (N51A) that retained CSR function but lost DNA deamination activity. In addition, an APOBEC1 mutation at N57, homologous to N51 of AID, also abolished DNA deamination activity but retained RNA editing activity. These results indicate that DNA deamination activity does not represent the physiological function of AID.
Conflict of interest statement
The authors declare no conflict of interest.
Figures



References
-
- Muramatsu M, et al. Specific expression of activation-induced cytidine deaminase (AID), a novel member of the RNA-editing deaminase family in germinal center B cells. J Biol Chem. 1999;274:18470–18476. - PubMed
-
- Revy P, et al. Activation-induced cytidine deaminase (AID) deficiency causes the autosomal recessive form of the Hyper-IgM syndrome (HIGM2) Cell. 2000;102:565–575. - PubMed
-
- Chaudhuri J, et al. Evolution of the immunoglobulin heavy chain class switch recombination mechanism. Adv Immunol. 2007;94:157–214. - PubMed
-
- Chaudhuri J, Tian M, Khuong C, Chua K, Pinaud E, Alt FW. Transcription-targeted DNA deamination by the AID antibody diversification enzyme. Nature. 2003;422:726–730. - PubMed
-
- Petersen-Mahrt SK, Harris RS, Neuberger MS. AID mutates E. coli suggesting a DNA deamination mechanism for antibody diversification. Nature. 2002;418:99–103. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
