

Subsets of myeloid-derived suppressor cells in tumor-bearing mice
- PMID: 18832739
- PMCID: PMC2575748
- DOI: 10.4049/jimmunol.181.8.5791
Subsets of myeloid-derived suppressor cells in tumor-bearing mice
Abstract
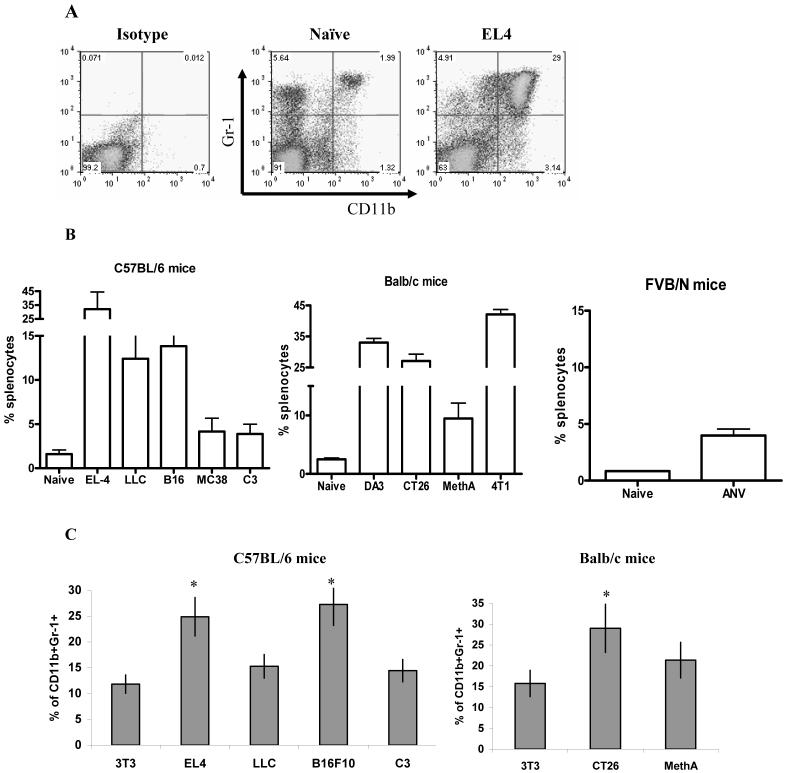
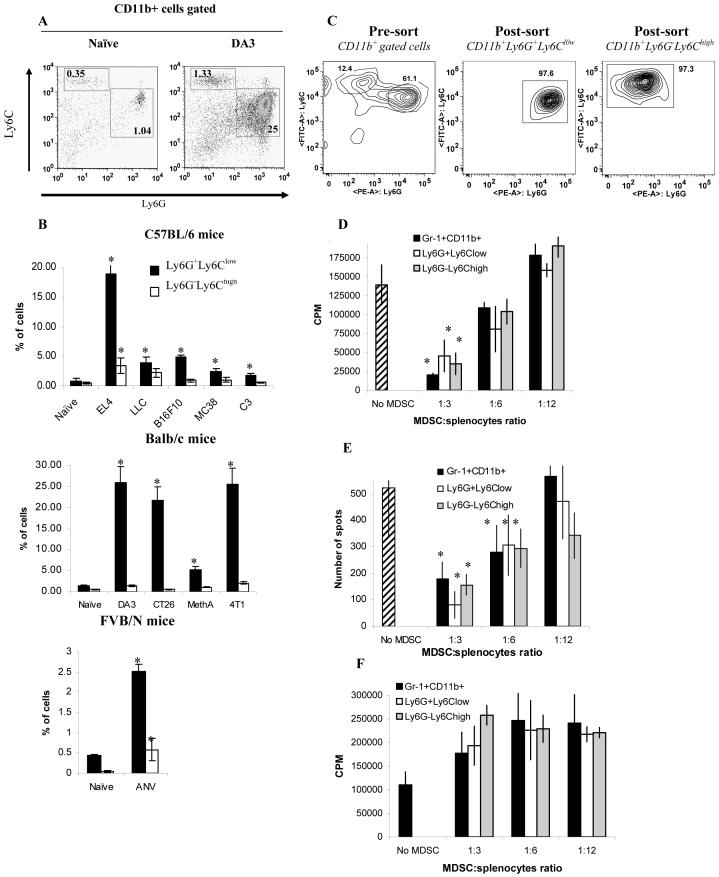
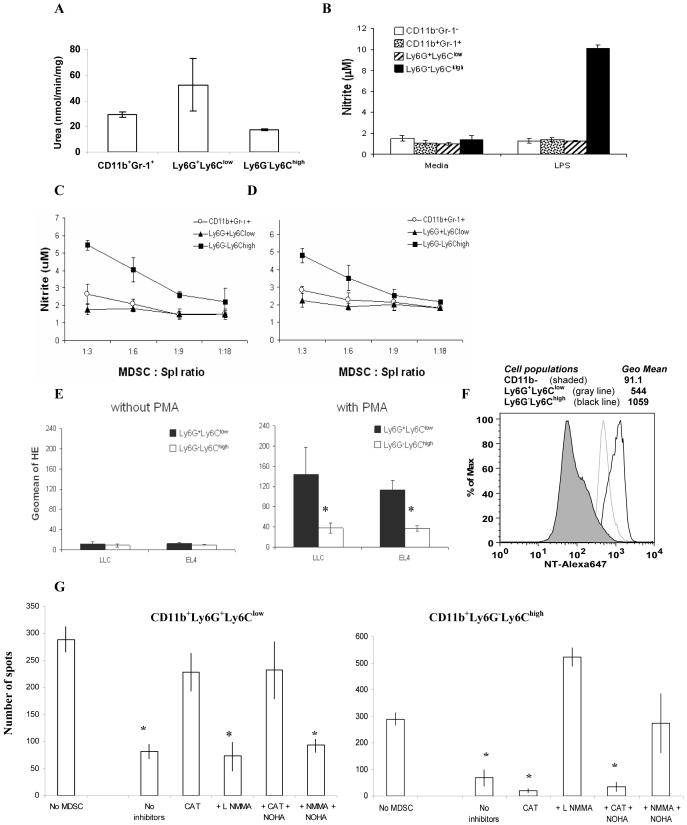
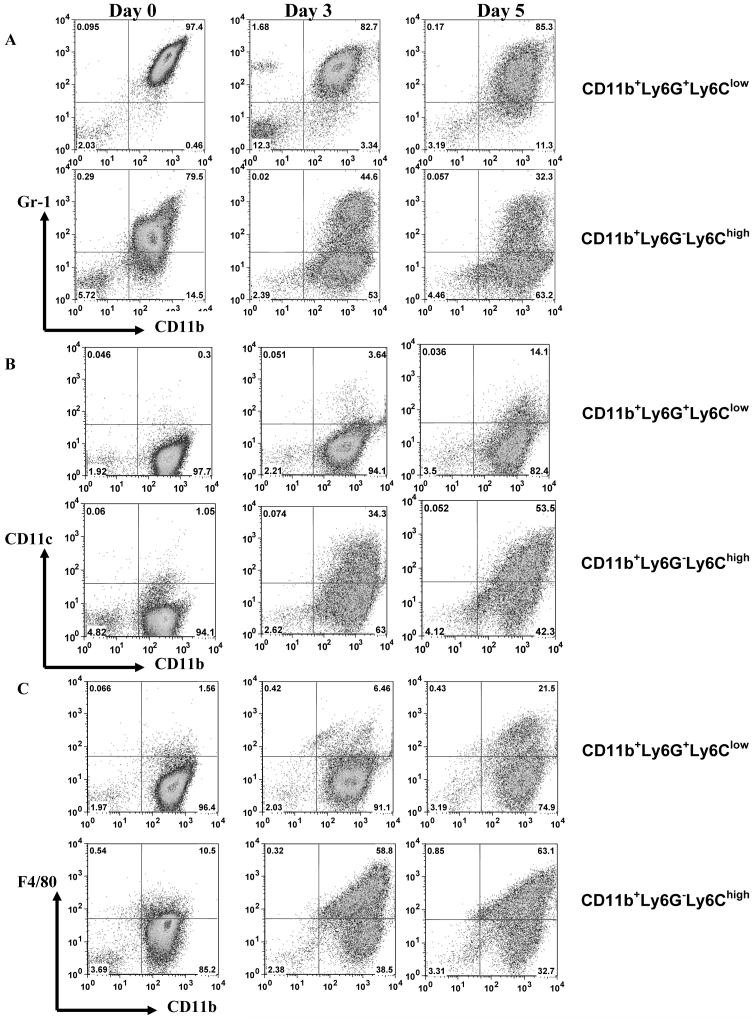
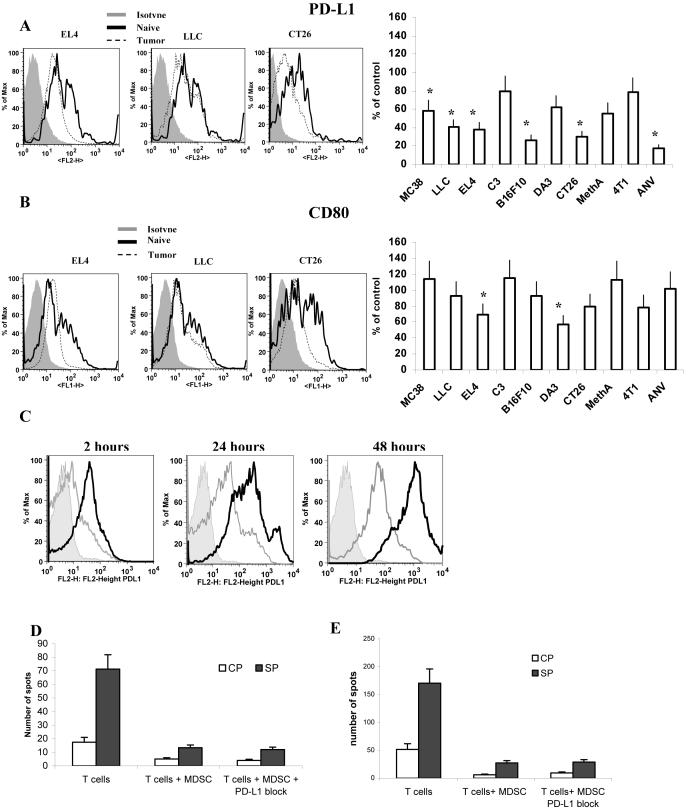
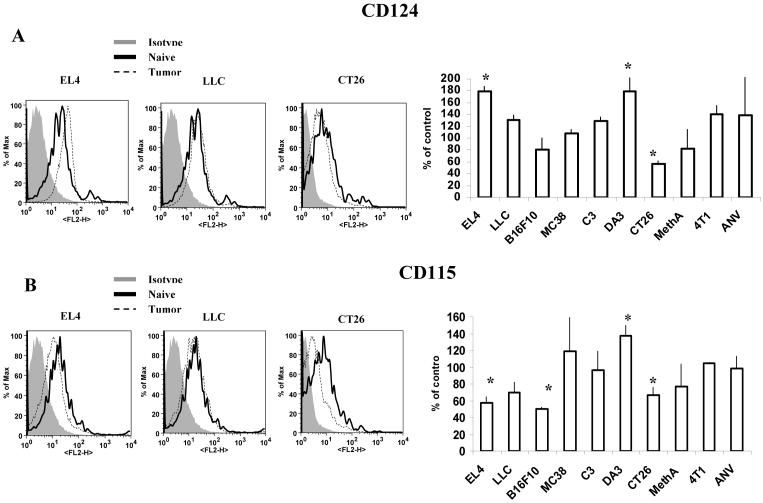
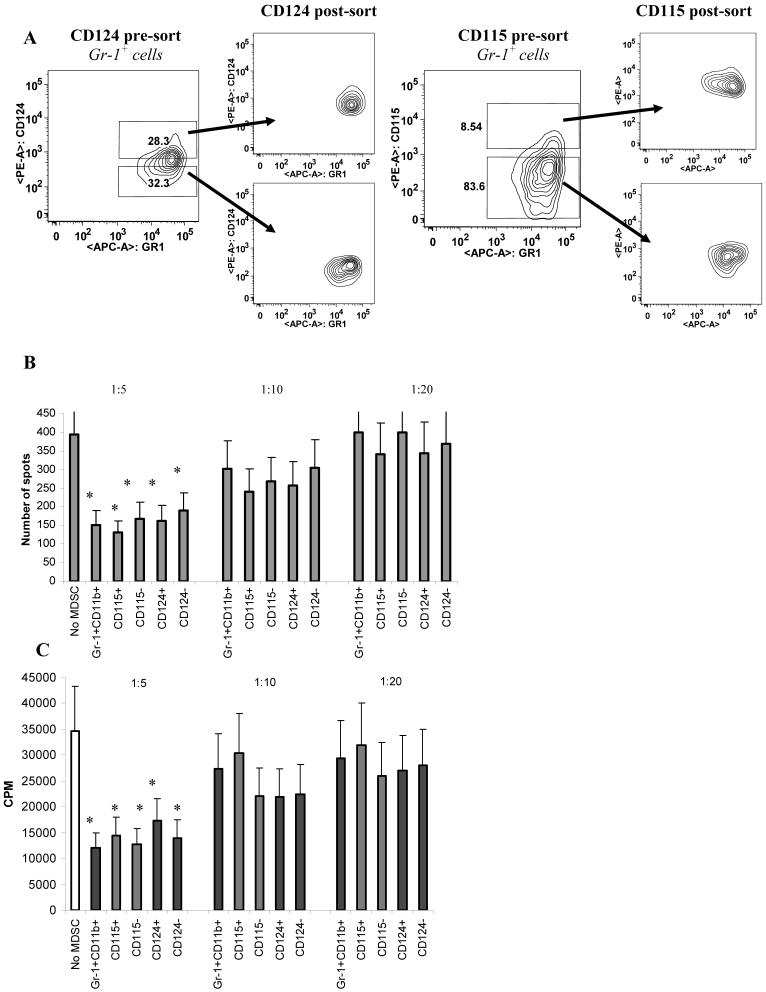
Myeloid-derived suppressor cells (MDSC) are a heterogeneous group of cells that play a critical role in tumor associated immune suppression. In an attempt to identify a specific subset of MDSC primarily responsible for immunosuppressive features of these cells, 10 different tumor models were investigated. All models showed variable but significant increase in the population of MDSC. Variability of MDSC expansion in vivo matched closely the effect of tumor cell condition medium in vitro. MDSC consists of two major subsets of Ly6G(+)Ly6C(low) granulocytic and Ly6G(-)Ly6C(high) monocytic cells. Granulocytic MDSC have increased level of reactive oxygen species and undetectable level of NO whereas monocytic MDSC had increased level of NO but undetectable levels of reactive oxygen species. However, their suppressive activity per cell basis was comparable. Almost all tumor models demonstrated a preferential expansion of granulocytic subset of MDSC. We performed a phenotypical and functional analysis of several surface molecules previously suggested to be involved in MDSC-mediated suppression of T cells: CD115, CD124, CD80, PD-L1, and PD-L2. Although substantial proportion of MDSC expressed those molecules no differences in the level of their expression or the proportion, positive cells were found between MDSC and cells from tumor-free mice that lack immune suppressive activity. The level of MDSC-mediated T cell suppression did not depend on the expression of these molecules. These data indicate that suppressive features of MDSC is caused not by expansion of a specific subset but more likely represent a functional state of these cells.
Figures
References
-
- Talmadge JE. Pathways mediating the expansion and immunosuppressive activity of myeloid-derived suppressor cells and their relevance to cancer therapy. Clin Cancer Res. 2007;13:5243–5248. - PubMed
-
- Delano MJ, Scumpia PO, Weinstein JS, Coco D, Nagaraj S, Kelly-Scumpia KM, O’Malley KA, Wynn JL, Antonenko S, Al-Quran SZ, Swan R, Chung CS, Atkinson MA, Ramphal R, Gabrilovich DI, Reeves WH, Ayala A, Phillips J, Laface D, Heyworth PG, Clare-Salzler M, Moldawer LL. MyD88-dependent expansion of an immature GR-1(+)CD11b(+) population induces T cell suppression and Th2 polarization in sepsis. J Exp Med. 2007;204:1463–1474. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
