

Hsp104, Hsp70 and Hsp40 interplay regulates formation, growth and elimination of Sup35 prions
- PMID: 18833196
- PMCID: PMC2572177
- DOI: 10.1038/emboj.2008.194
Hsp104, Hsp70 and Hsp40 interplay regulates formation, growth and elimination of Sup35 prions
Abstract
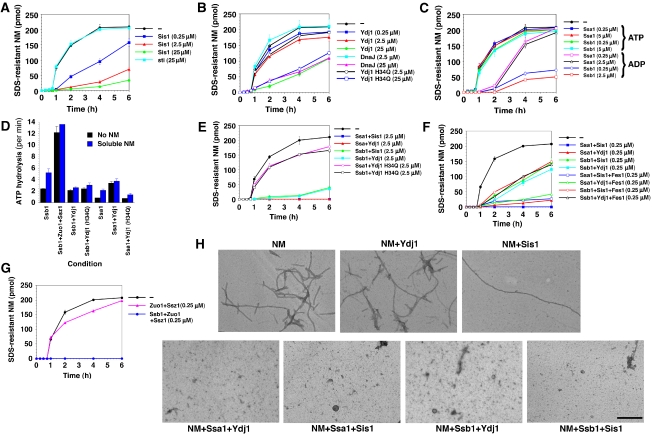
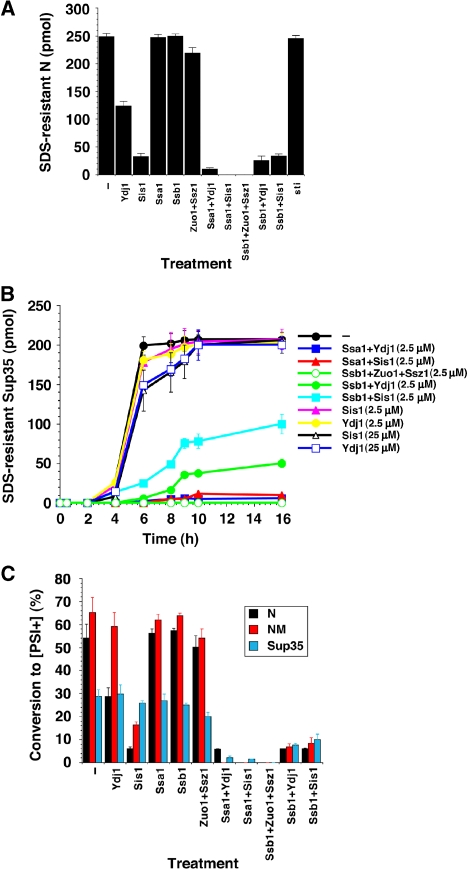
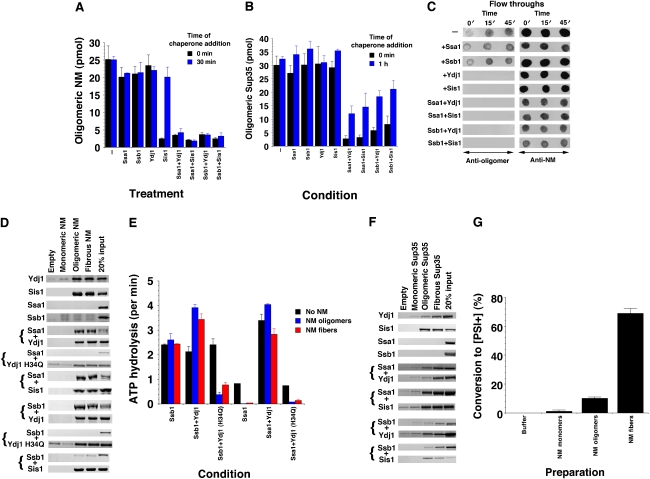
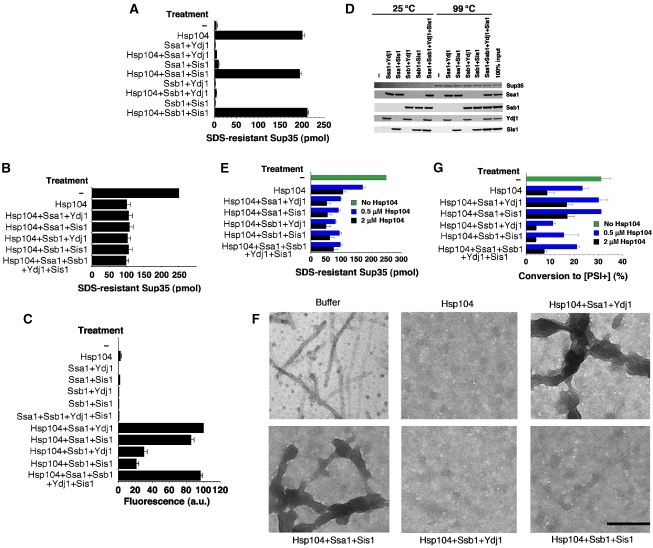
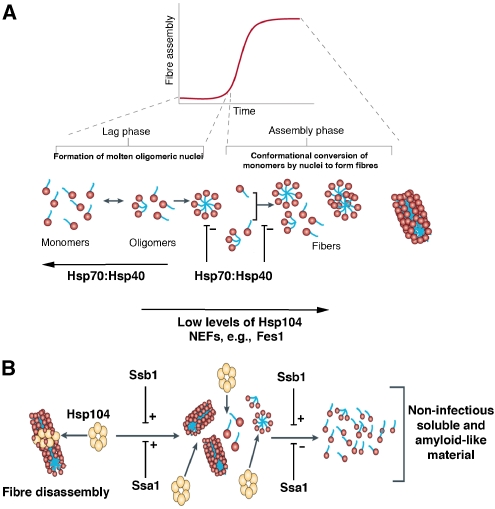
Self-templating amyloid forms of Sup35 constitute the yeast prion [PSI(+)]. How the protein-remodelling factor, Hsp104, collaborates with other chaperones to regulate [PSI(+)] inheritance remains poorly delineated. Here, we report how the Ssa and Ssb components of the Hsp70 chaperone system directly affect Sup35 prionogenesis and cooperate with Hsp104. We identify the ribosome-associated Ssb1:Zuo1:Ssz1 complex as a potent antagonist of Sup35 prionogenesis. The Hsp40 chaperones, Sis1 and Ydj1, preferentially interact with Sup35 oligomers and fibres compared with monomers, and facilitate Ssa1 and Ssb1 binding. Various Hsp70:Hsp40 pairs block prion nucleation by disassembling molten oligomers and binding mature oligomers. By binding fibres, Hsp70:Hsp40 pairs occlude prion recognition elements and inhibit seeded assembly. These inhibitory activities are partially relieved by the nucleotide exchange factor, Fes1. Low levels of Hsp104 stimulate prionogenesis and alleviate inhibition by some Hsp70:Hsp40 pairs. At high concentrations, Hsp104 eliminates Sup35 prions. This activity is reduced when Ssa1, or enhanced when Ssb1, is incorporated into nascent prions. These findings illuminate several facets of the chaperone interplay that underpins [PSI(+)] inheritance.
Figures


References
-
- Albanese V, Yam AY, Baughman J, Parnot C, Frydman J (2006) Systems analyses reveal two chaperone networks with distinct functions in eukaryotic cells. Cell 124: 75–88 - PubMed
-
- Chernoff YO, Lindquist SL, Ono B, Inge-Vechtomov SG, Liebman SW (1995) Role of the chaperone protein Hsp104 in propagation of the yeast prion-like factor [PSI+]. Science 268: 880–884 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
