

Visualizing transient events in amino-terminal autoprocessing of HIV-1 protease
- PMID: 18833280
- PMCID: PMC2798589
- DOI: 10.1038/nature07342
Visualizing transient events in amino-terminal autoprocessing of HIV-1 protease
Abstract
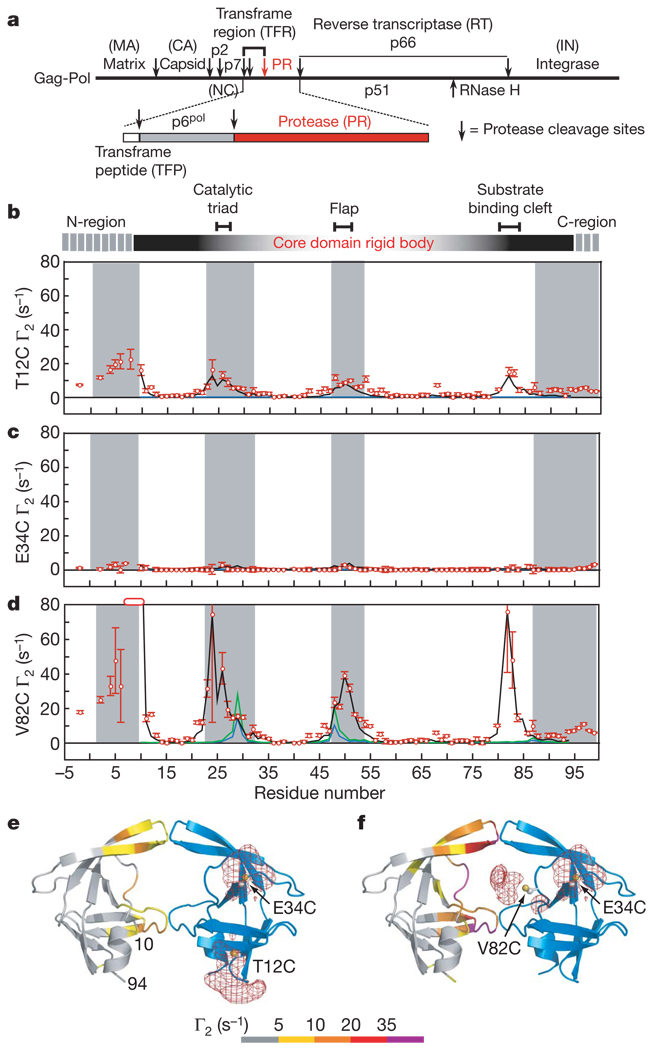
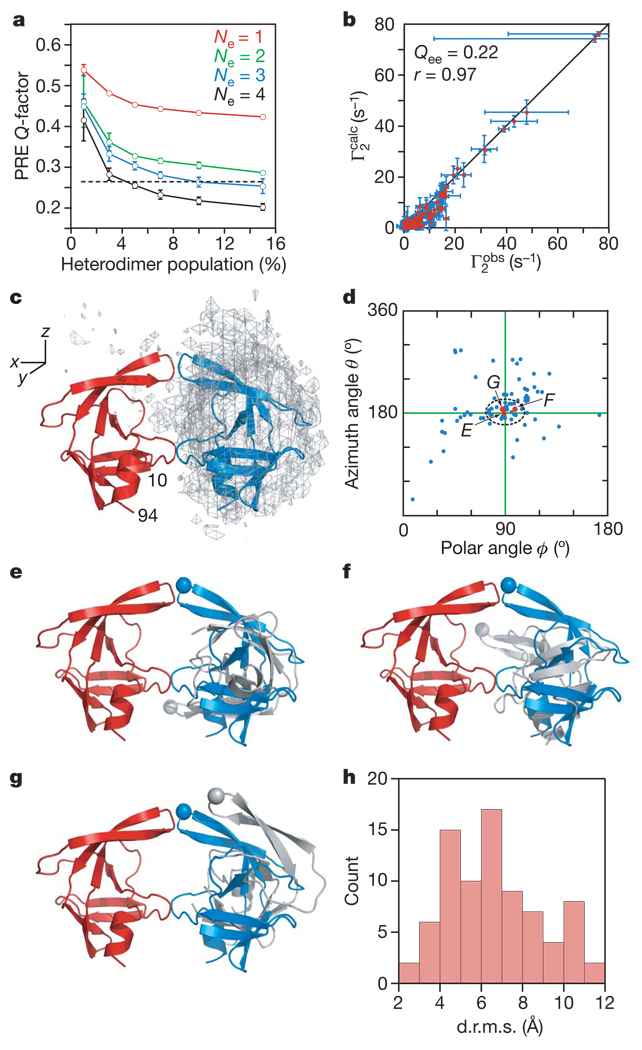
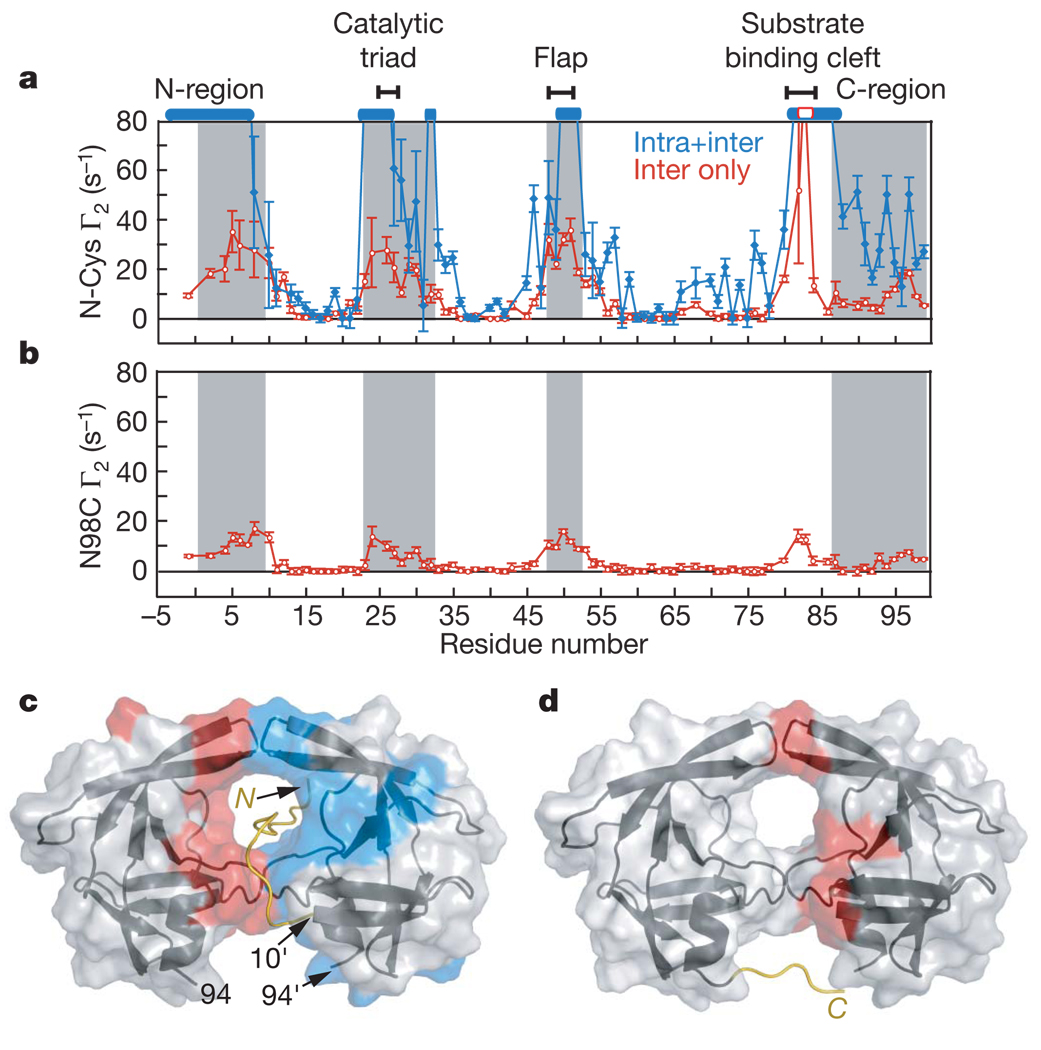
HIV-1 protease processes the Gag and Gag-Pol polyproteins into mature structural and functional proteins, including itself, and is therefore indispensable for viral maturation. The mature protease is active only as a dimer with each subunit contributing catalytic residues. The full-length transframe region protease precursor appears to be monomeric yet undergoes maturation via intramolecular cleavage of a putative precursor dimer, concomitant with the appearance of mature-like catalytic activity. How such intramolecular cleavage can occur when the amino and carboxy termini of the mature protease are part of an intersubunit beta-sheet located distal from the active site is unclear. Here we visualize the early events in N-terminal autoprocessing using an inactive mini-precursor with a four-residue N-terminal extension that mimics the transframe region protease precursor. Using paramagnetic relaxation enhancement, a technique that is exquisitely sensitive to the presence of minor species, we show that the mini-precursor forms highly transient, lowly populated (3-5%) dimeric encounter complexes that involve the mature dimer interface but occupy a wide range of subunit orientations relative to the mature dimer. Furthermore, the occupancy of the mature dimer configuration constitutes a very small fraction of the self-associated species (accounting for the very low enzymatic activity of the protease precursor), and the N-terminal extension makes transient intra- and intersubunit contacts with the substrate binding site and is therefore available for autocleavage when the correct dimer orientation is sampled within the encounter complex ensemble.
Figures
References
-
- Louis JM, Weber IT, Tozser J, Clore GM, Gronenborn AM. HIV-1 protease: maturation, enzyme specificity, and drug resistance. Adv. Pharmacol. 2000;49:111–146. - PubMed
-
- Louis JM, Ishima R, Torchia DA, Weber IT. HIV-1 protease: structure, dynamics and inhibition. Adv. Pharmacol. 2007;55:261–298. - PubMed
-
- Wlodawer A, Erikson J. Structure-based inhibitors of HIV-1 protease. Annu. Rev. Biochem. 1993;62:543–585. - PubMed
-
- Wlodawer A, Vondrasek J. Inhibitors of HIV-1 protease: a major success of structure-assisted drug design. Annu. Rev. Biophys. Biomol. Struct. 1998;27:249–284. - PubMed
-
- Ishima R, Torchia DA, Lynch SM, Gronenborn AM, Louis JM. Solution structure of the mature HIV-1 protease monomer: insight into the tertiary fold and stability of a precursor. J. Biol. Chem. 2003;278:43311–43319. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
