

Anti cancer effects of curcumin: cycle of life and death
- PMID: 18834508
- PMCID: PMC2572158
- DOI: 10.1186/1747-1028-3-14
Anti cancer effects of curcumin: cycle of life and death
Abstract
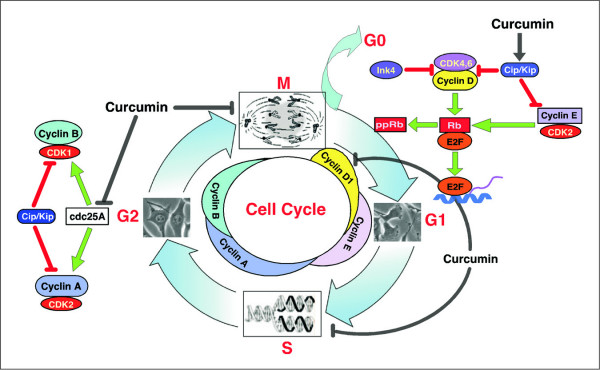
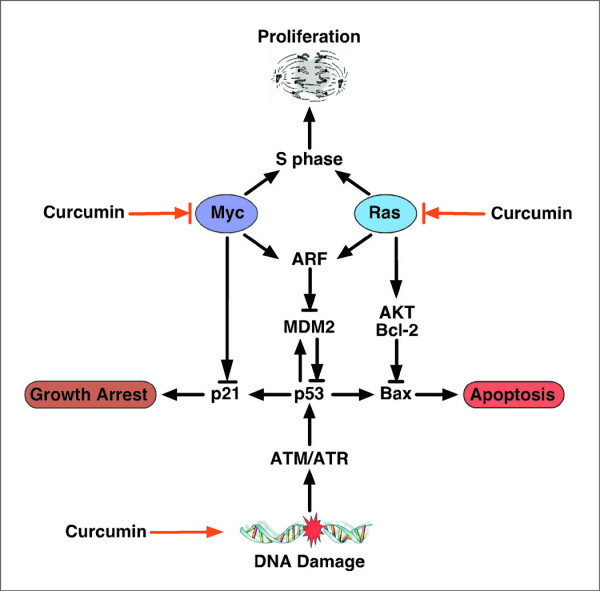
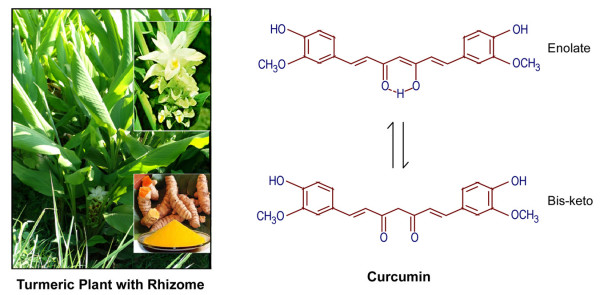
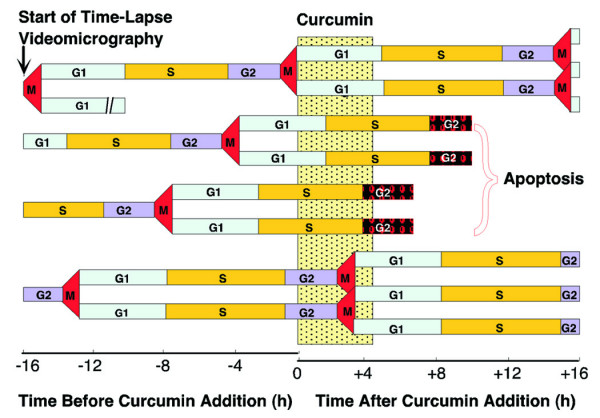
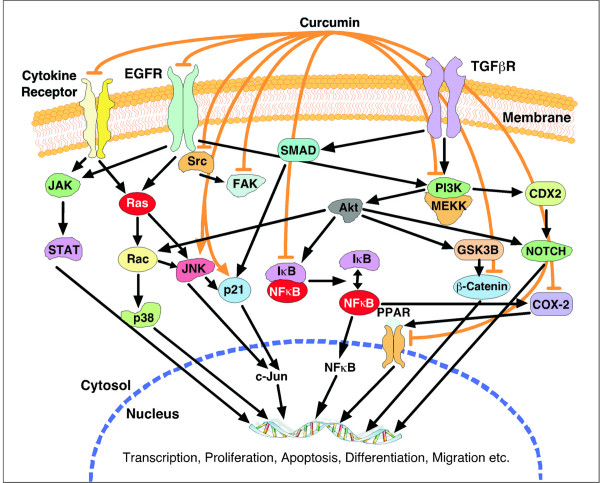
Increasing knowledge on the cell cycle deregulations in cancers has promoted the introduction of phytochemicals, which can either modulate signaling pathways leading to cell cycle regulation or directly alter cell cycle regulatory molecules, in cancer therapy. Most human malignancies are driven by chromosomal translocations or other genetic alterations that directly affect the function of critical cell cycle proteins such as cyclins as well as tumor suppressors, e.g., p53. In this respect, cell cycle regulation and its modulation by curcumin are gaining widespread attention in recent years. Extensive research has addressed the chemotherapeutic potential of curcumin (diferuloylmethane), a relatively non-toxic plant derived polyphenol. The mechanisms implicated are diverse and appear to involve a combination of cell signaling pathways at multiple levels. In the present review we discuss how alterations in the cell cycle control contribute to the malignant transformation and provide an overview of how curcumin targets cell cycle regulatory molecules to assert anti-proliferative and/or apoptotic effects in cancer cells. The purpose of the current article is to present an appraisal of the current level of knowledge regarding the potential of curcumin as an agent for the chemoprevention of cancer via an understanding of its mechanism of action at the level of cell cycle regulation. Taken together, this review seeks to summarize the unique properties of curcumin that may be exploited for successful clinical cancer prevention.
Figures
References
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
