

Rapid regulation of telomere length is mediated by poly(ADP-ribose) polymerase-1
- PMID: 18835851
- PMCID: PMC2577345
- DOI: 10.1093/nar/gkn615
Rapid regulation of telomere length is mediated by poly(ADP-ribose) polymerase-1
Abstract
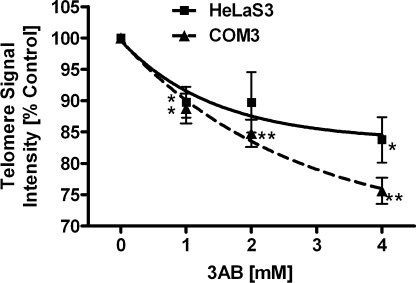
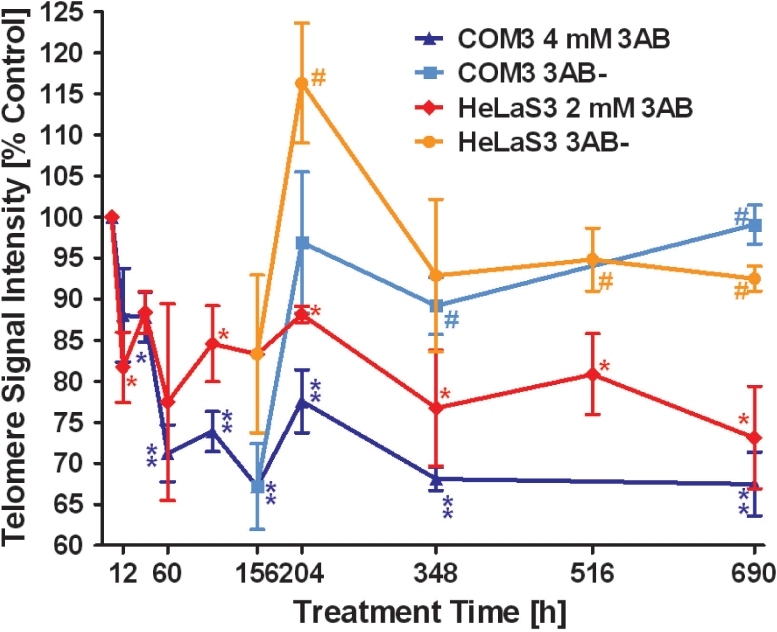
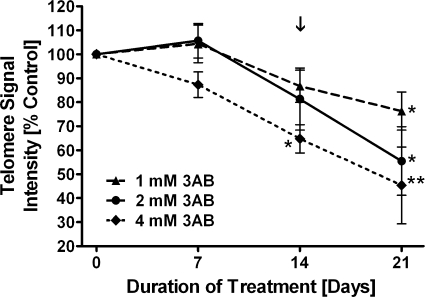
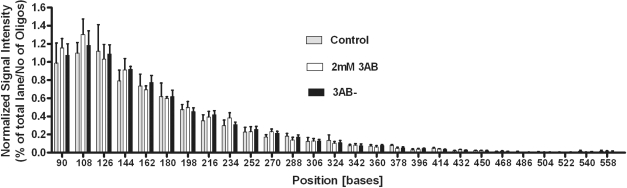
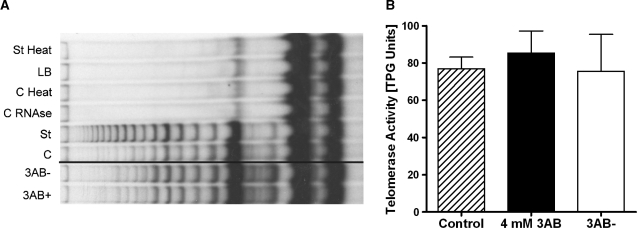
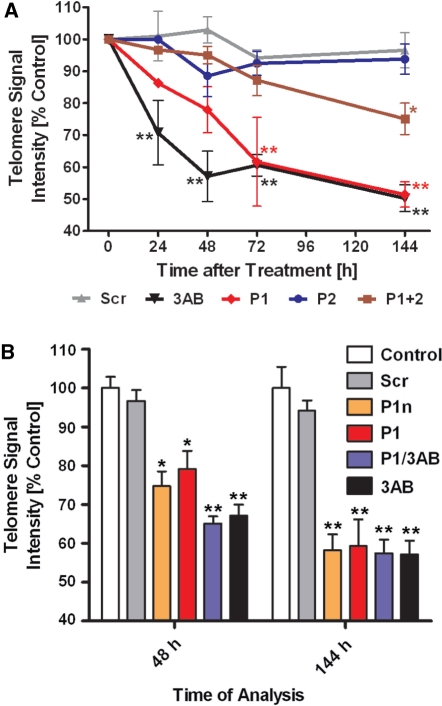
Shelterin/telosome is a multi-protein complex at mammalian telomeres, anchored to the double-stranded region by the telomeric-repeat binding factors-1 and -2. In vitro modification of these proteins by poly(ADP-ribosyl)ation through poly(ADP-ribose) polymerases-5 (tankyrases) and -1/-2, respectively, impairs binding. Thereafter, at least telomeric-repeat binding factor-1 is degraded by the proteasome. We show that pharmacological inhibition of poly(ADP-ribose) polymerase activity in cells from two different species leads to rapid decrease in median telomere length and stabilization at a lower setting. Specific knockdown of poly(ADP-ribose) polymerase-1 by RNA interference had the same effect. The length of the single-stranded telomeric overhang as well as telomerase activity were not affected. Release of inhibition led to a fast re-gain in telomere length to control levels in cells expressing active telomerase. We conclude that poly(ADP-ribose) polymerase-1 activity and probably its interplay with telomeric-repeat binding factor-2 is an important determinant in telomere regulation. Our findings reinforce the link between poly(ADP-ribosyl)ation and aging/longevity and also impact on the use of poly(ADP-ribose) polymerase inhibitors in tumor therapy.
Figures
References
-
- Celli GB, de Lange T. DNA processing is not required for ATM-mediated telomere damage response after TRF2 deletion. Nat. Cell Biol. 2005;7:712–718. - PubMed
-
- Liu D, O’Connor MS, Qin J, Songyang Z. Telosome, a mammalian telomere-associated complex formed by multiple telomeric proteins. J. Biol. Chem. 2004;279:51338–51342. - PubMed
-
- Griffith JD, Comeau L, Rosenfield S, Stansel RM, Bianchi A, Moss H, de Lange T. Mammalian telomeres end in a large duplex loop. Cell. 1999;97:503–514. - PubMed
-
- Smith S, Giriat I, Schmitt A, de Lange T. Tankyrase, a poly(ADP-ribose) polymerase at human telomeres. Science. 1998;282:1484–1487. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
