

Nuclear receptor co-regulator Krüppel-like factor 9 and prohibitin 2 expression in estrogen-induced epithelial cell proliferation in the mouse uterus
- PMID: 18835980
- PMCID: PMC2612732
- DOI: 10.1677/JOE-08-0383
Nuclear receptor co-regulator Krüppel-like factor 9 and prohibitin 2 expression in estrogen-induced epithelial cell proliferation in the mouse uterus
Abstract
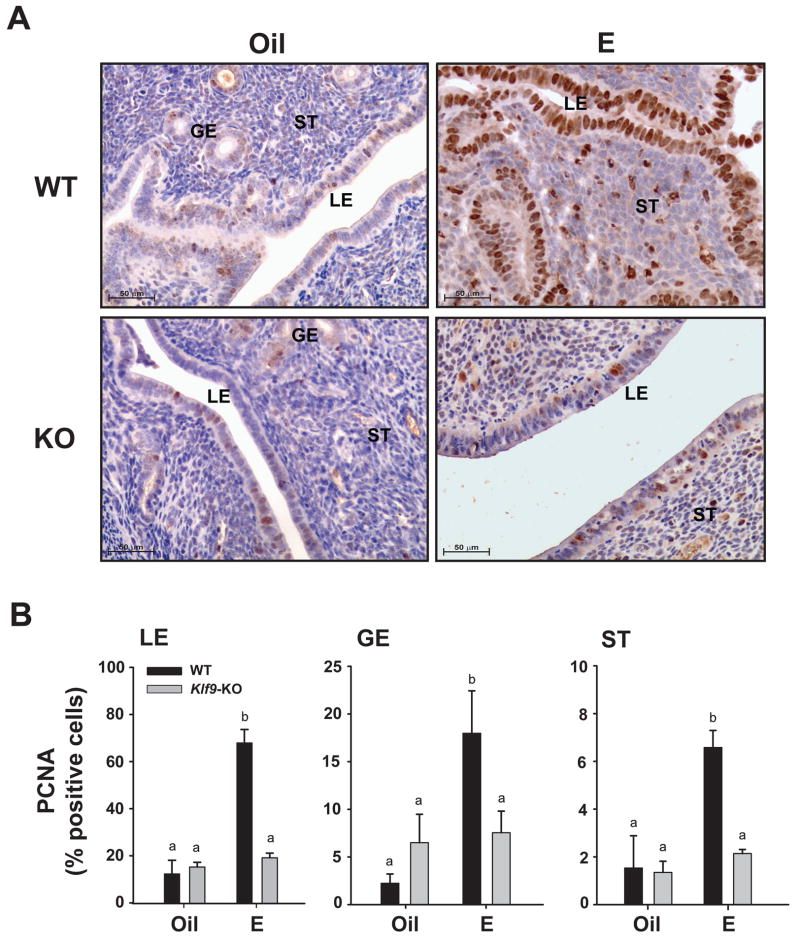
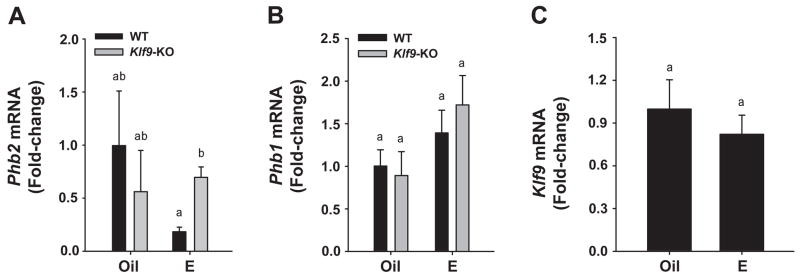
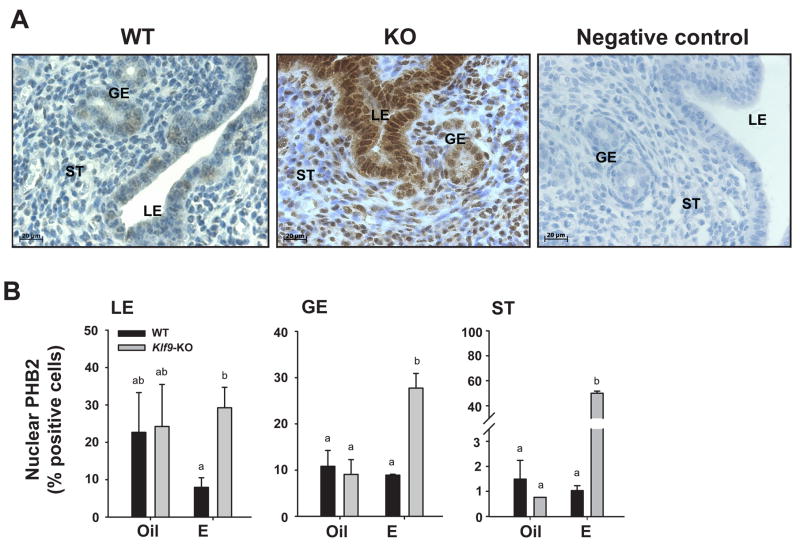
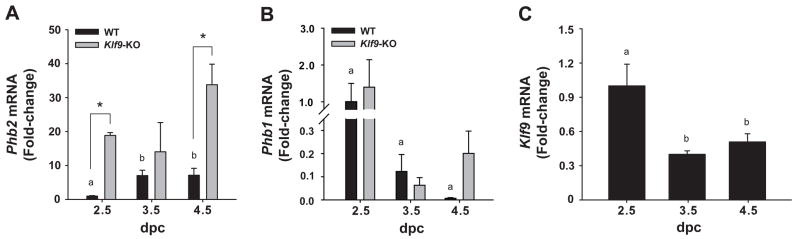
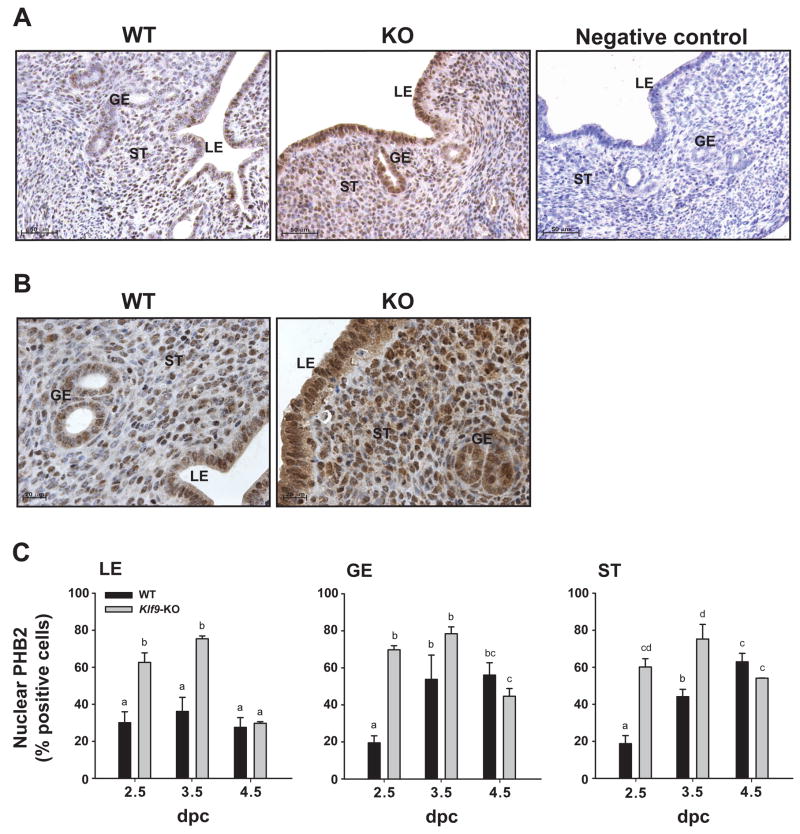
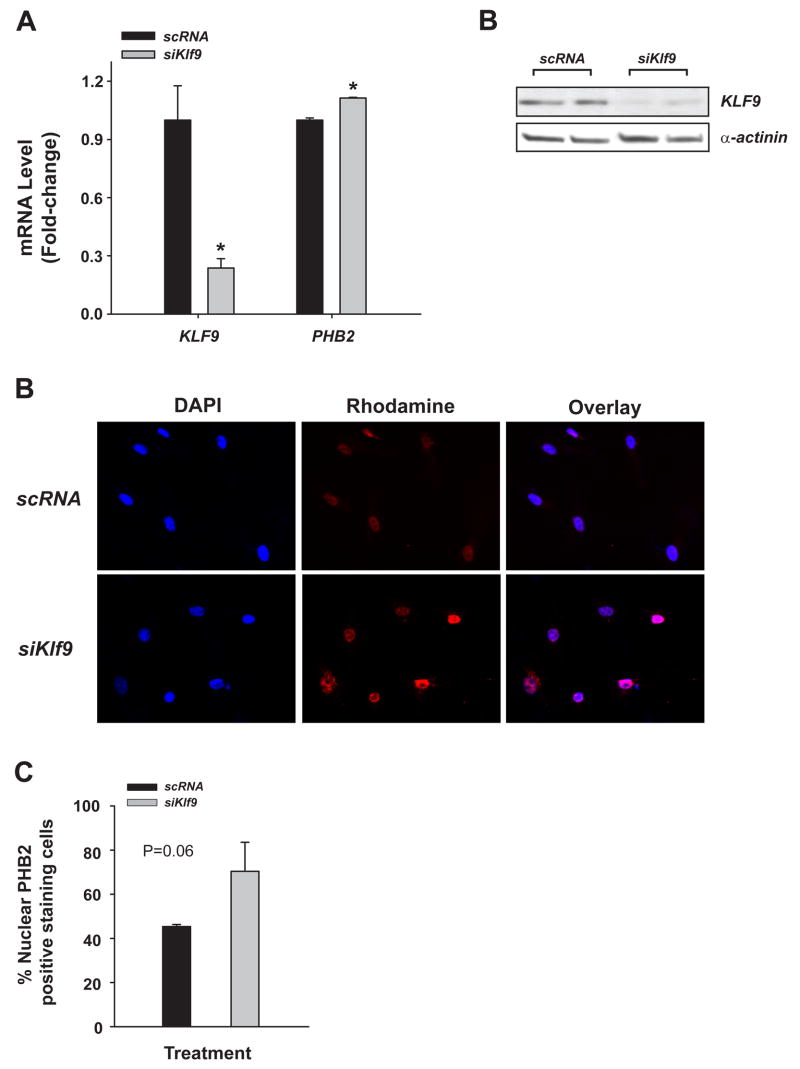
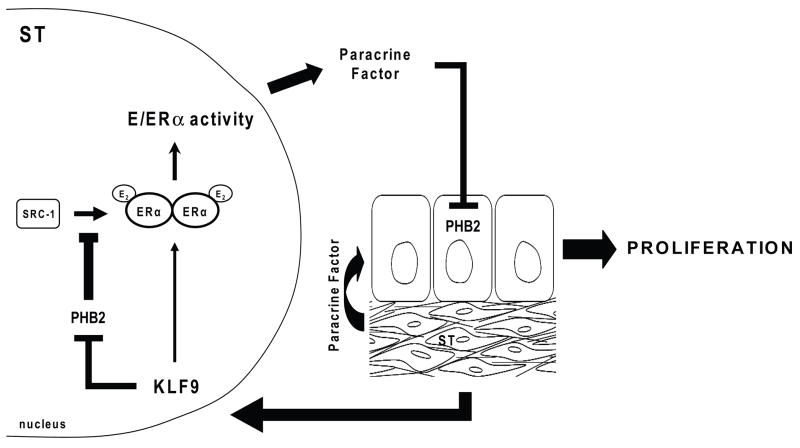
Estrogen, acting through its cognate receptor estrogen receptor-alpha (ESR1), is a critical regulator of uterine endometrial epithelial proliferation. Although the dynamic communication between endometrial stromal (ST) and epithelial cells is considered to be an important component in this process, key molecular players in particular compartments remain poorly defined. Here, we used mice null for Krüppel-like factor 9 (KLF9) to evaluate the contribution of this nuclear protein in ST-epithelial interactions underlying proliferative effects of estrogen. We found that in ovariectomized mice administered estradiol-17beta (E(2)) for 24 h, Klf9 null mutation resulted in lack of E(2)-induced proliferative response in all endometrial compartments. We demonstrated a negative association between Klf9 expression and nuclear levels of ESR1 transcriptional corepressor prohibitin (PHB) 2 in uterine ST and epithelial cells of E(2)-treated wild-type (WT) and Klf9 null mice. In early pregnancy uteri of WT mice, the temporal pattern of Klf9 transcript levels was inversely associated with that of Phb2. Deletion of Klf9 up-regulated uterine Phb2 expression and increased PHB2 nuclear localization in endometrial ST and epithelial cells, with no effects on the expression of the related Phb1. In the human endometrial ST cell line treated with E(2) for 24 h, Klf9 siRNA targeting augmented PHB2 transcript and increased nuclear PHB2 protein levels, albeit this effect was not to the extent seen in vivo with Klf9 null mutants. Our findings suggest a novel mechanism for control of estrogen-induced luminal epithelial proliferation involving ST KLF9 regulation of paracrine factor(s) to repress epithelial expression of corepressor PHB2.
Figures
References
-
- Delage-Morroux R, Martini PG, Choi I, Kraichely DM, Hoeksema J, Katzenellenbogen BS. Analysis of estrogen receptor interaction with repressor of estrogen receptor activity (REA) and the regulation of estrogen receptor transcriptional activity by REA. J Biol Chem. 2000;275:35848–35856. - PubMed
-
- Dupont S, Krust A, Gansmuller A, Dierich A, Chambon P, Mark M. Effect of single and compound knockouts of estrogen receptors alpha (ERalpha) and beta (ERbeta) on mouse reproductive phenotypes. Development. 2000;127:4277–4291. - PubMed
-
- Dutertre M, Smith CL. Ligand-independent interactions of p160/steroid receptor coactivators and CREB-binding protein (CBP) with estrogen receptor α: regulation by phosphorylation sites in the A/B region depends on other receptor domains. Mol Endocrinol. 2003;17:1296–1314. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
