

E2F in vivo binding specificity: comparison of consensus versus nonconsensus binding sites
- PMID: 18836037
- PMCID: PMC2577861
- DOI: 10.1101/gr.080622.108
E2F in vivo binding specificity: comparison of consensus versus nonconsensus binding sites
Abstract
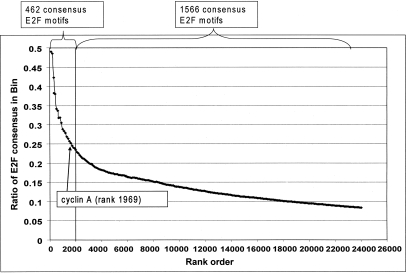
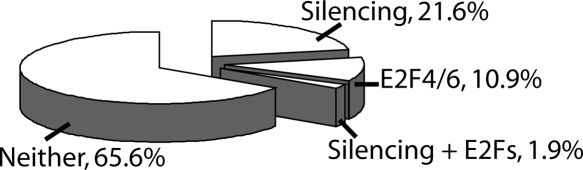
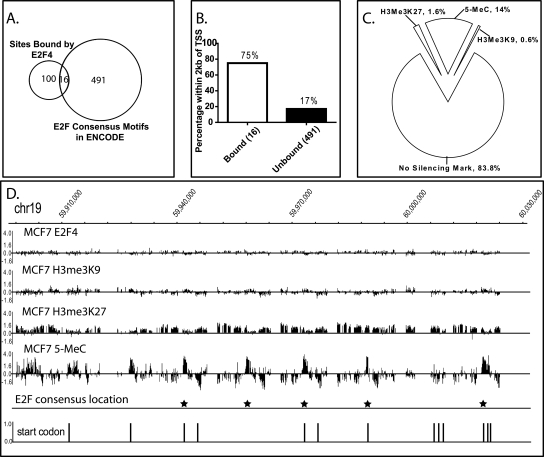
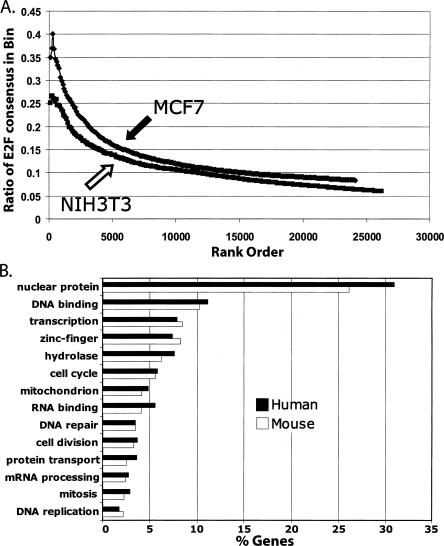
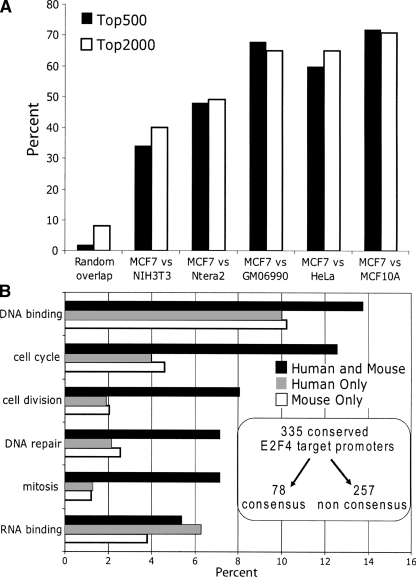
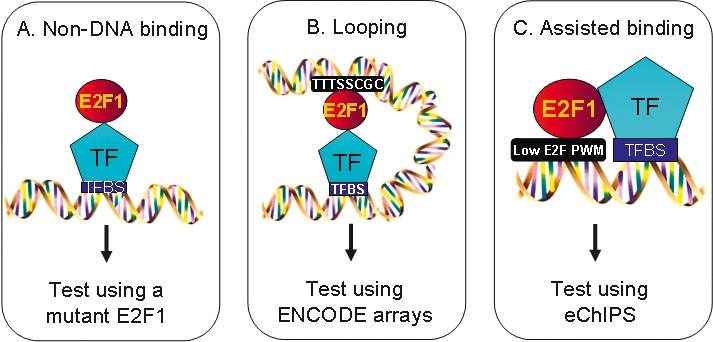
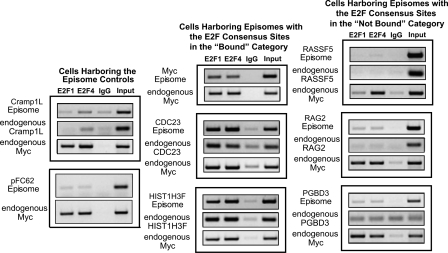
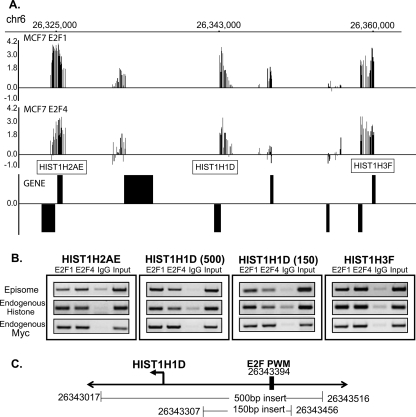
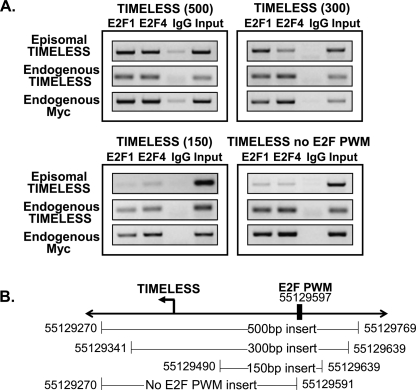
We have previously shown that most sites bound by E2F family members in vivo do not contain E2F consensus motifs. However, differences between in vivo target sites that contain or lack a consensus E2F motif have not been explored. To understand how E2F binding specificity is achieved in vivo, we have addressed how E2F family members are recruited to core promoter regions that lack a consensus motif and are excluded from other regions that contain a consensus motif. Using chromatin immunoprecipitation coupled with DNA microarray analysis (ChIP-chip) assays, we have shown that the predominant factors specifying whether E2F is recruited to an in vivo binding site are (1) the site must be in a core promoter and (2) the region must be utilized as a promoter in that cell type. We have tested three models for recruitment of E2F to core promoters lacking a consensus site, including (1) indirect recruitment, (2) looping to the core promoter mediated by an E2F bound to a distal motif, and (3) assisted binding of E2F to a site that weakly resembles an E2F motif. To test these models, we developed a new in vivo assay, termed eChIP, which allows analysis of transcription factor binding to isolated fragments. Our findings suggest that in vivo (1) a consensus motif is not sufficient to recruit E2Fs, (2) E2Fs can bind to isolated regions that lack a consensus motif, and (3) binding can require regions other than the best match to the E2F motif.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases