

A neural code for three-dimensional object shape in macaque inferotemporal cortex
- PMID: 18836443
- PMCID: PMC2725445
- DOI: 10.1038/nn.2202
A neural code for three-dimensional object shape in macaque inferotemporal cortex
Abstract
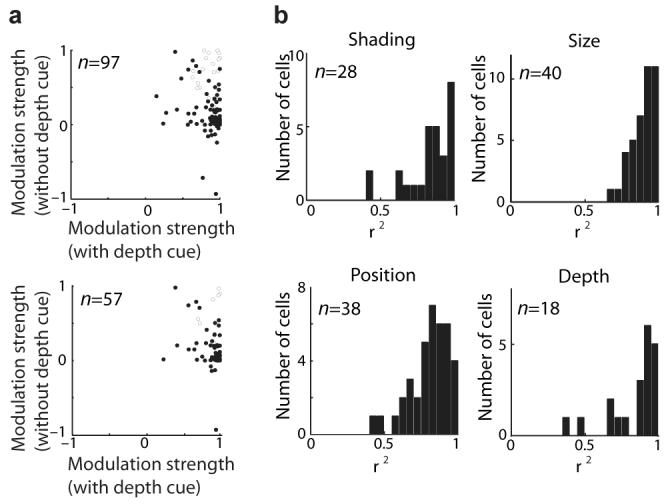
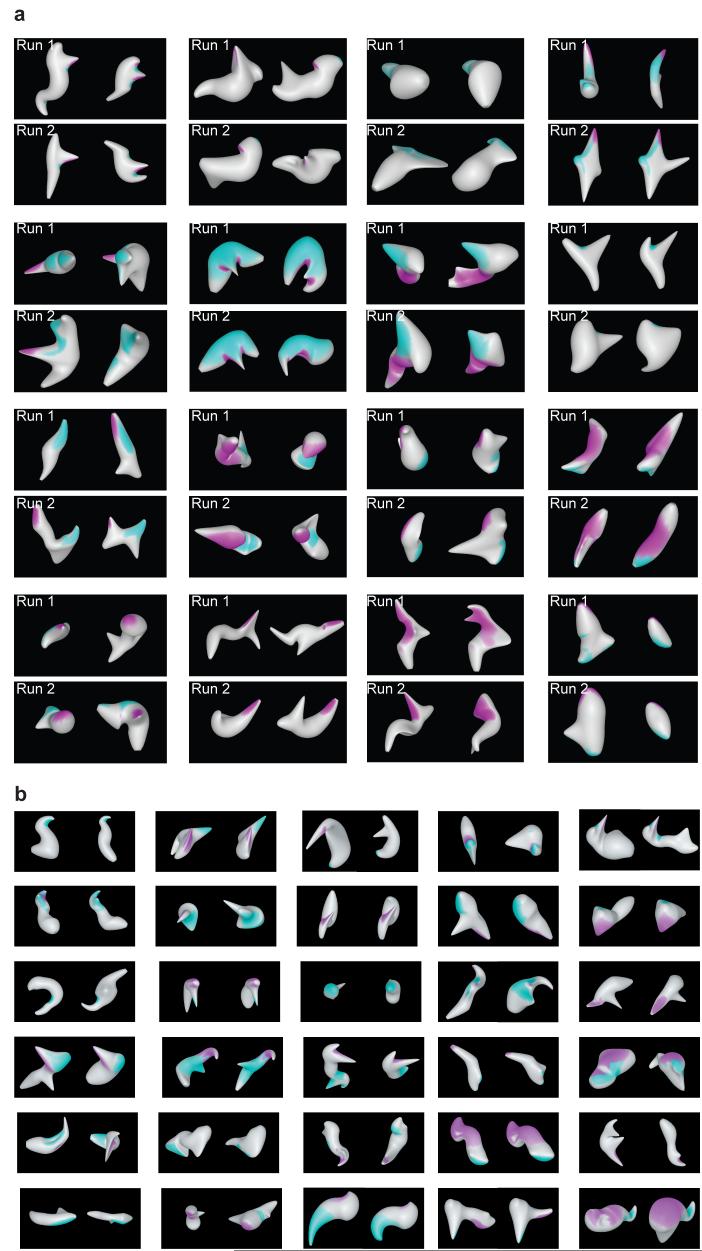
Previous investigations of the neural code for complex object shape have focused on two-dimensional pattern representation. This may be the primary mode for object vision given its simplicity and direct relation to the retinal image. In contrast, three-dimensional shape representation requires higher-dimensional coding derived from extensive computation. We found evidence for an explicit neural code for complex three-dimensional object shape. We used an evolutionary stimulus strategy and linear/nonlinear response models to characterize three-dimensional shape responses in macaque monkey inferotemporal cortex (IT). We found widespread tuning for three-dimensional spatial configurations of surface fragments characterized by their three-dimensional orientations and joint principal curvatures. Configural representation of three-dimensional shape could provide specific knowledge of object structure to support guidance of complex physical interactions and evaluation of object functionality and utility.
Figures




Comment in
-
So many pixels, so little time.Nat Neurosci. 2008 Nov;11(11):1243-4. doi: 10.1038/nn1108-1243. Nat Neurosci. 2008. PMID: 18956009 No abstract available.
References
-
- Ungerleider LG, Mishkin M. In: Analysis of Visual Behavior. Ingle DG, Goodale MA, Mansfield RJQ, editors. MIT Press; Cambridge, Massachusetts: 1982. pp. 549–586.
-
- Felleman DJ, Van Essen DC. Distributed hierarchical processing in the primate cerebral cortex. Cereb. Cortex. 1991;1:1–47. - PubMed
-
- Anzai A, Peng X, Van Essen DC. Neurons in monkey visual area V2 encode combinations of orientations. Nat. Neurosci. 2007;10:1313–1321. - PubMed
-
- Gallant JL, Braun J, Van Essen DC. Selectivity for polar, hyperbolic, and Cartesian gratings in macaque visual cortex. Science. 1993;259:100–103. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
