

Review
doi: 10.1016/j.mce.2008.09.005.
Epub 2008 Sep 17.
The aging male hypothalamic-pituitary-gonadal axis: pulsatility and feedback
Affiliations
- PMID: 18838102
- PMCID: PMC2662347
- DOI: 10.1016/j.mce.2008.09.005
Item in Clipboard
Review
The aging male hypothalamic-pituitary-gonadal axis: pulsatility and feedback
Mol Cell Endocrinol.
.
Abstract
Aging results in insidious decremental changes in hypothalamic, pituitary and gonadal function. The foregoing three main anatomic loci of control are regulated by intermittent time-delayed signal exchange, principally via gonadotropin-releasing hormone (GnRH), luteinizing hormone (LH) and testosterone/estradiol (Te/E(2)). A mathematical framework is required to embody these dynamics. The present review highlights integrative adaptations in the aging male hypothalamic-pituitary-gonadal axis, as assessed by recent objective ensemble models of the axis as a whole.
Figures
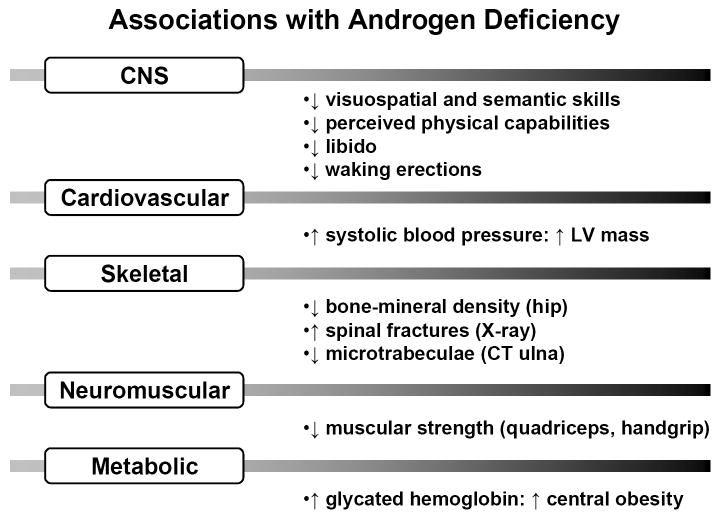
Epidemiological associations reported with androgen deficiency in men. CNS = central nervous system; LV = left ventricular; CT = computed tomography

Decline of total, bioavailable and free Te concentrations with age in healthy men in Olmsted County, MN (unpublished cross-sectional data).
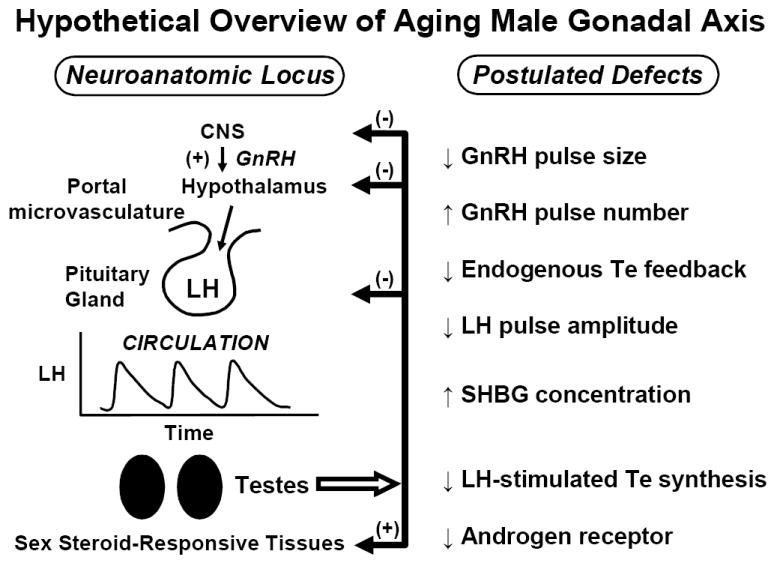
Hypothesized deficits in of aging human male gonadal axis deduced from indirect evidence. Smaller, more frequent GnRH pulses might occur under reduced negative feedback by systemic Te (or E2) resulting in low-amplitude high-frequency LH pulses. The efficacy of LH in stimulating Te secretion also appears to decline in aging individuals. Whether Te’s actions on the brain, pituitary gland and prostate via the androgen receptor (AR) are diminished is not established. Unpublished line drawing.
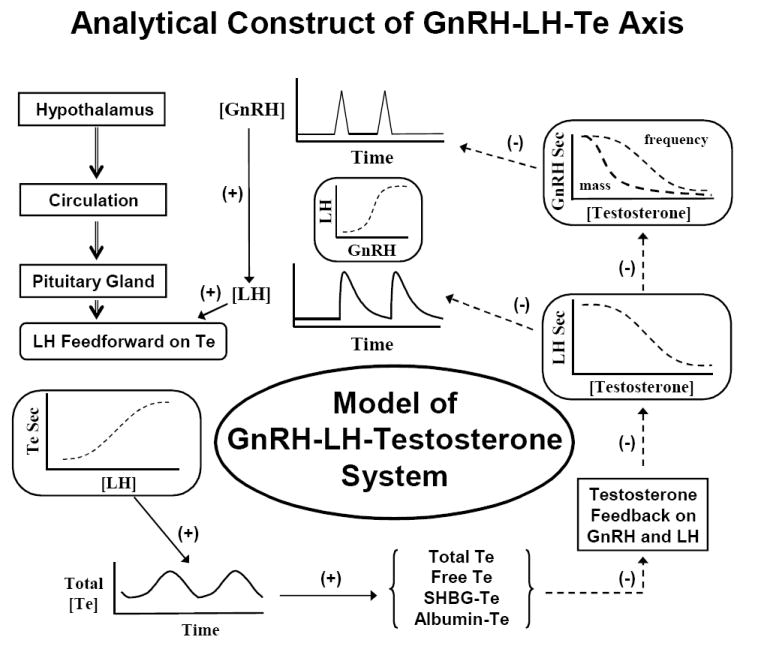
Ensemble model of GnRH, LH and testosterone (Te) signals, which are mathematically linked by nonlinear stimulatory (feedforward) and inhibitory (feedback) dose-response functions and time delays. In this ensemble structure, the hypothalamus, pituitary, gland, testis and circulation constitute anatomic loci, which communicate dynamically via intermittent signaling. Signals, such as GnRH, LH and Te, act on target sites via nonlinear (sigmoidal) dose-response functions. A goal is to estimate such functions noninvasively in vivo in aging individuals. In principle, individual or combined dose-response effects could deteriorate in aging. Adapted from (Keenan et al., 2004).
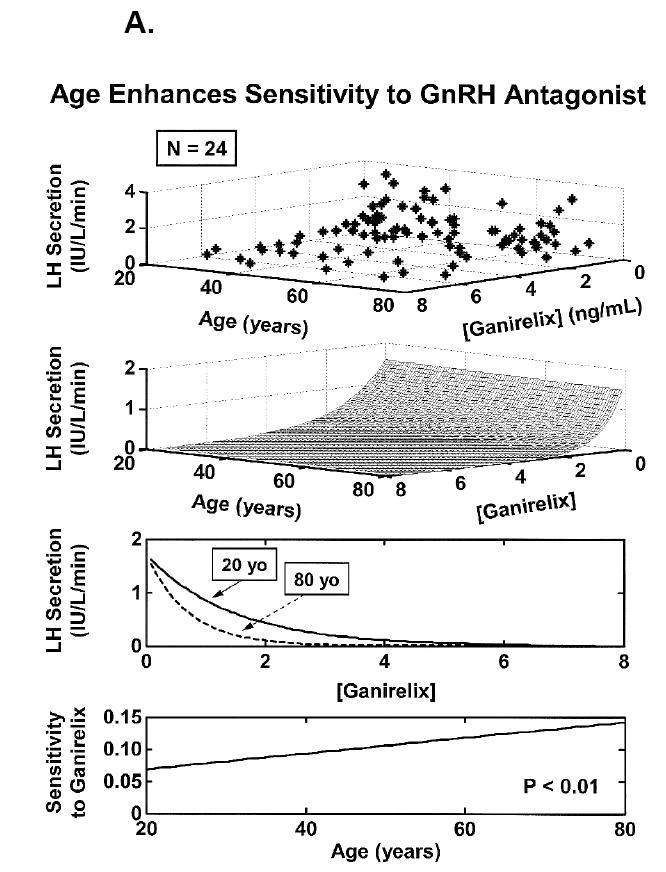
Concomitant decline of endogenous GnRH outflow and efficacy of LH drive with age. Panel A. Analytical reconstruction of LH secretion (vertical axis) as a joint function of GnRH-receptor antagonist (ganirelix) concentrations (horizontal axis) and age (oblique axis). Greater sensitivity of LH secretion to ganirelix inhibition in older men forecasts less opposition by endogenous GnRH drive. Panel B. Analogous 3-dimensional surface plot for LH → Te feedforward dose-response curves as a function of age (top). Data are from 24 healthy men ages 20 to 72 yr each studied under a 4-strata ganirelix clamp. Boxed curves are extrapolated by regression (bottom). Adapted from (Keenan et al., 2006).
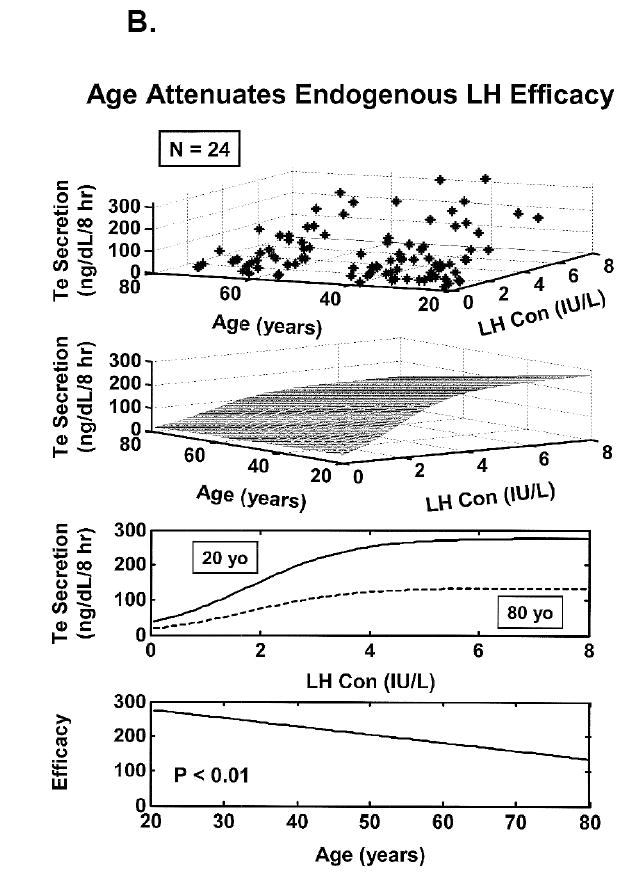
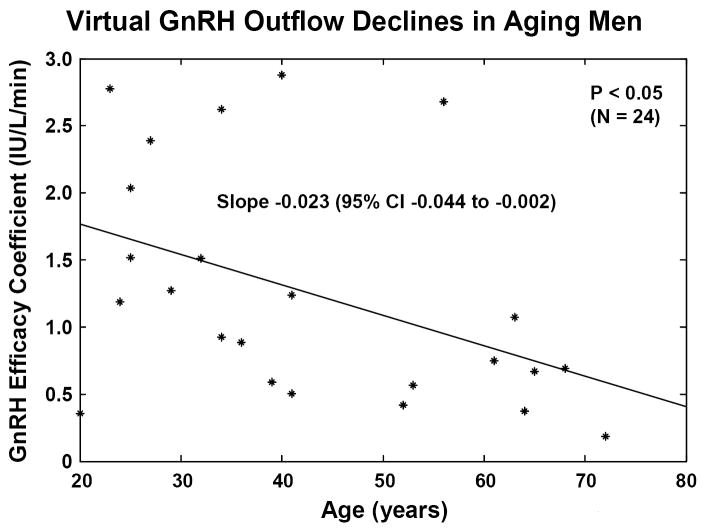
Concomitant decline of endogenous GnRH outflow and efficacy of LH drive with age. Panel A. Analytical reconstruction of LH secretion (vertical axis) as a joint function of GnRH-receptor antagonist (ganirelix) concentrations (horizontal axis) and age (oblique axis). Greater sensitivity of LH secretion to ganirelix inhibition in older men forecasts less opposition by endogenous GnRH drive. Panel B. Analogous 3-dimensional surface plot for LH → Te feedforward dose-response curves as a function of age (top). Data are from 24 healthy men ages 20 to 72 yr each studied under a 4-strata ganirelix clamp. Boxed curves are extrapolated by regression (bottom). Adapted from (Keenan et al., 2006).
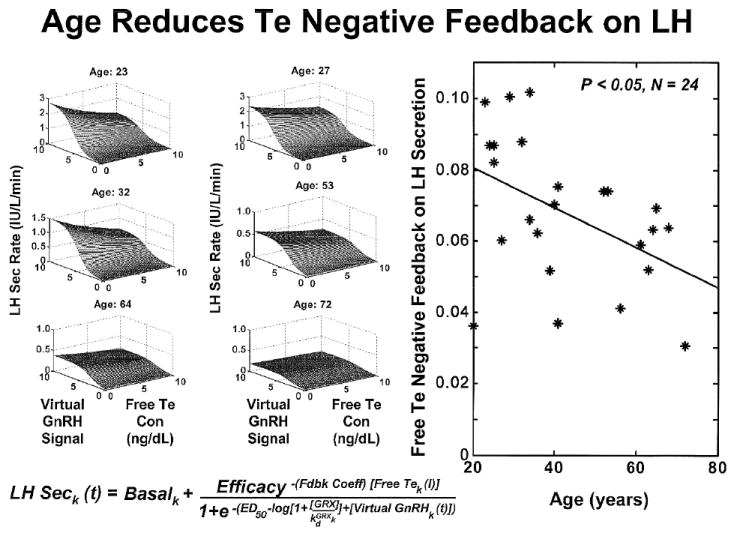
Noninvasive analytical reconstruction of joint feedforward (GnRH concentrations → LH secretion) and feedback (Te concentrations → GnRH/LH secretion) response surfaces in 6 of 24 healthy men of the indicated ages. The vertical (z-axis) projection of the surface is determined jointly by GnRH outflow and free Te feedback. Impaired feedback is inferred by a lesser drop in LH secretion rates as free Te concentrations increase between 0 and 10 ng/dL at any given GnRH level in older subjects. Adapted from data in (Keenan et al., 2006).
Analytical estimation of feedforward by GnRH concentrations onto LH secretion rates [defined as efficacy of GnRH drive to gonadotropes] in 24 healthy men ages 20-72 yr. Adapted from data in (Keenan et al., 2006).

Preservation (top, eugonadal placebo) (bottom, low-Te clamp) of GnRH dose-response properties in older (age > 60 yr) compared with young (age < 35 yr) healthy men. GnRH doses are in ng/kg. Each GnRH pulse was injected on a separate randomly ordered morning fasting. Data are the mean ± SEM. Adapted from (Veldhuis et al., 2005a).
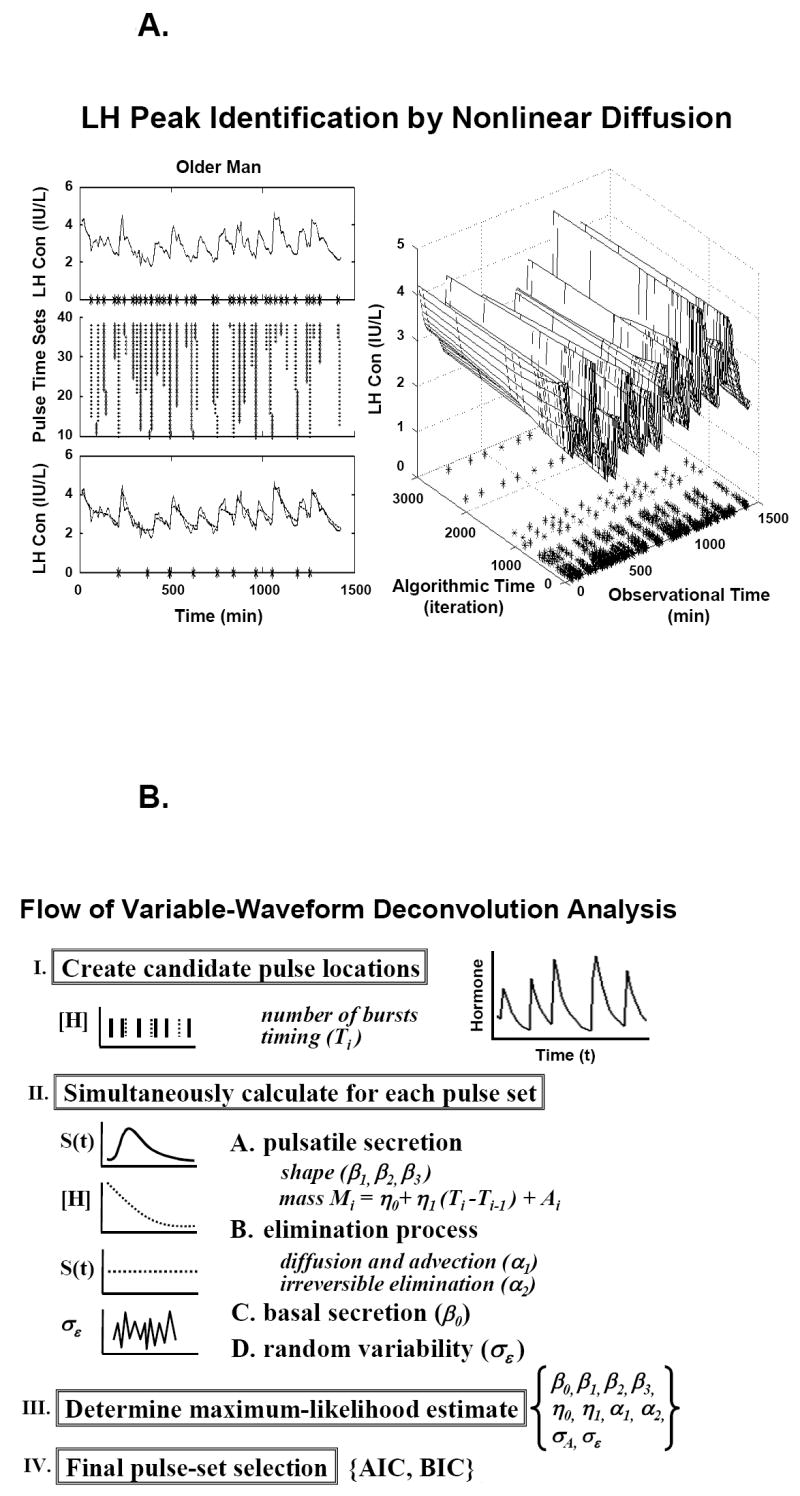
Panel A. Schema of pulse-detection phase of a third-generation flexible (Gamma) waveform deconvolution method. Asterisks denote pulse-onset times (observational time) retained after repeated incremental smoothing (algorithmic time). Panel B. Quantification of secretory-burst number, size and shape, basal secretion, biexponential elimination, stochastic variability in pulse mass, and experimental uncertainty in the data conditioned on a set of a priori candidate pulse-onset times [Panel A]. Greek symbols define parameters. AIC = Akaike and BIC = Bayesian information criteria are used to select the final pulse set. Unpublished line drawings from methods presented in (Keenan et al., 2003; Keenan et al., 2005; Keenan et al., 2006; Keenan et al., 2004).
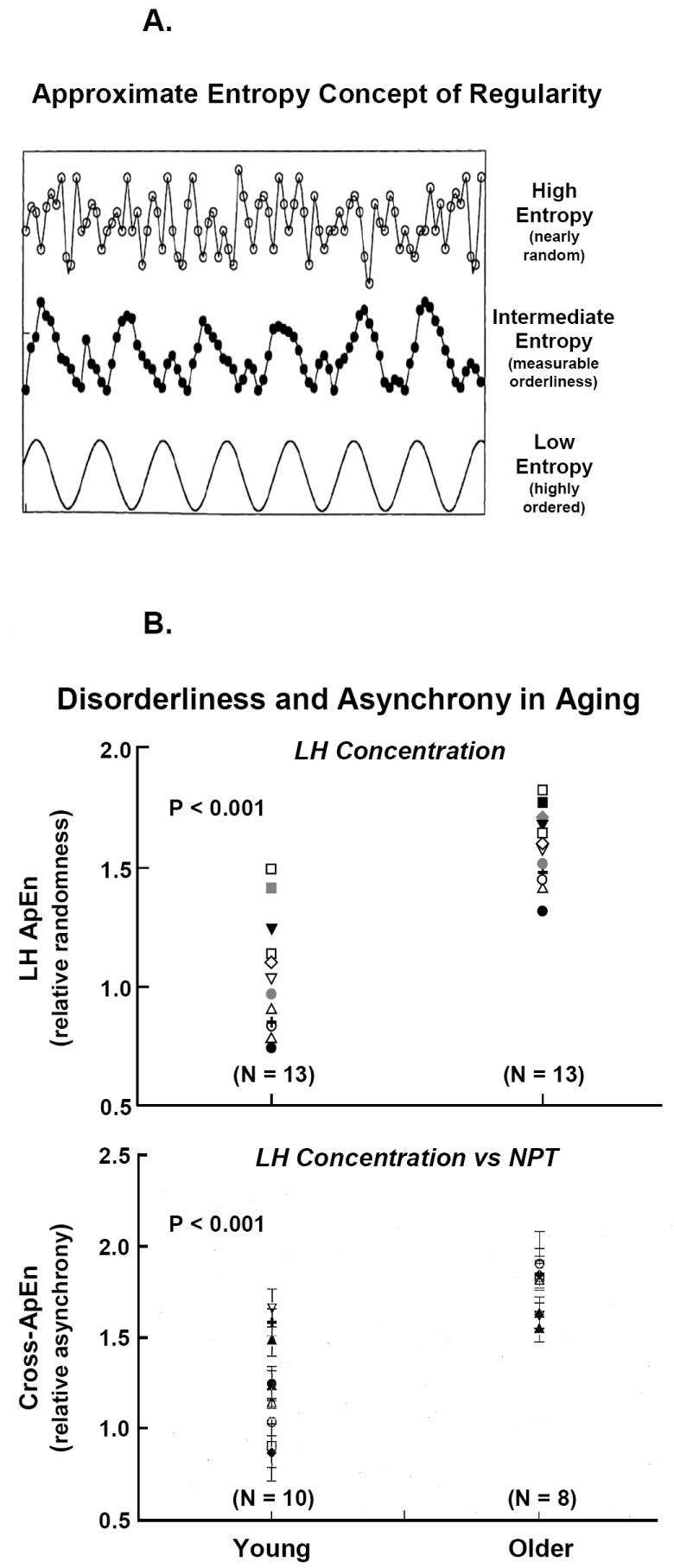
Panel A. Concept of approximate entropy (ApEn) to quantify the orderliness or regularity of hormone concentration or secretion patterns. Orderliness is a measure of feedback control in coupled systems. The bottom curve gives a cosine function (low ApEn or well ordered), the middle plot less regular data, and the top frame an irregular profile (high ApEn). Panel B. Elevated LH ApEn (top) in individual older compared with young men signifies reduced negative feedback on the secretion process. Elevated cross-ApEn (bottom) of LH outflow and nocturnal penile tumescence (NPT) oscillations denotes loss of neurohormone-outflow synchrony in aging individuals. Data are from (Veldhuis et al., 1999).
Similar articles
-
Hypothalamic-pituitary-gonadal axis in two men with aromatase deficiency: evidence that circulating estrogens are required at the hypothalamic level for the integrity of gonadotropin negative feedback.Eur J Endocrinol. 2006 Oct;155(4):513-22. doi: 10.1530/eje.1.02254. Eur J Endocrinol. 2006. PMID: 16990650
-
Regulation of complex pulsatile and rhythmic neuroendocrine systems: the male gonadal axis as a prototype.Prog Brain Res. 2010;181:79-110. doi: 10.1016/S0079-6123(08)81006-0. Prog Brain Res. 2010. PMID: 20478434 Review.
-
Two-week pulsatile gonadotropin releasing hormone infusion unmasks dual (hypothalamic and Leydig cell) defects in the healthy aging male gonadotropic axis.Eur J Endocrinol. 1999 Sep;141(3):257-66. doi: 10.1530/eje.0.1410257. Eur J Endocrinol. 1999. PMID: 10474123 Clinical Trial.
-
Central aspects of systemic oestradiol negative- and positive-feedback on the reproductive neuroendocrine system.J Neuroendocrinol. 2020 Jan;32(1):e12724. doi: 10.1111/jne.12724. Epub 2019 May 23. J Neuroendocrinol. 2020. PMID: 31054210 Free PMC article. Review.
-
Effects of aging on the hypothalamic-hypophyseal-gonadal axis in female rats.Fertil Steril. 1977 Dec;28(12):1365-70. doi: 10.1016/s0015-0282(16)42986-9. Fertil Steril. 1977. PMID: 201506
Cited by
-
Why men age faster but reproduce longer than women: mTOR and evolutionary perspectives.Aging (Albany NY). 2010 May;2(5):265-73. doi: 10.18632/aging.100149. Aging (Albany NY). 2010. PMID: 20519781 Free PMC article.
-
Improved early postnatal nutrition and its effect on histomorphological parameters in the testes of Sanjabi ram lambs.Trop Anim Health Prod. 2019 Jul;51(6):1539-1544. doi: 10.1007/s11250-019-01842-0. Epub 2019 Feb 13. Trop Anim Health Prod. 2019. PMID: 30759296
-
Feedback on LH in Testosterone-Clamped Men Depends on the Mode of Testosterone Administration and Body Composition.J Endocr Soc. 2018 Nov 23;3(1):235-249. doi: 10.1210/js.2018-00317. eCollection 2019 Jan 1. J Endocr Soc. 2018. PMID: 30623162 Free PMC article.
-
Short-Acting Testosterone: More Physiologic?Front Endocrinol (Lausanne). 2020 Sep 30;11:572465. doi: 10.3389/fendo.2020.572465. eCollection 2020. Front Endocrinol (Lausanne). 2020. PMID: 33117287 Free PMC article. No abstract available.
-
Effects of Heat Stress-Induced Sex Hormone Dysregulation on Reproduction and Growth in Male Adolescents and Beneficial Foods.Nutrients. 2024 Sep 8;16(17):3032. doi: 10.3390/nu16173032. Nutrients. 2024. PMID: 39275346 Free PMC article. Review.
References
-
- Akingbemi BT, Ge R, Rosenfeld CS, Newton LG, Hardy DO, Catterall JF, Lubahn DB, Korach KS, Hardy MP. Estrogen receptor-alpha gene deficiency enhances androgen biosynthesis in the mouse Leydig cell. Endocrinol. 2003;144:84–93. - PubMed
-
- Cao L, Leers-Sucheta S, Azhar S. Aging alters the functional expression of enzymatic and non-enzymatic anti-oxidant defense systems in testicular rat Leydig cells. J Steroid Biochem Mol Biol. 2004;88:61–67. - PubMed
-
- Clarke IJ, Cummins JT. The significance of small pulses of gonadotropin-releasing hormone. J Endocrinol. 1987;113:413–418. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
