

Conformational changes involved in G-protein-coupled-receptor activation
- PMID: 18838178
- PMCID: PMC3475190
- DOI: 10.1016/j.tips.2008.08.006
Conformational changes involved in G-protein-coupled-receptor activation
Abstract
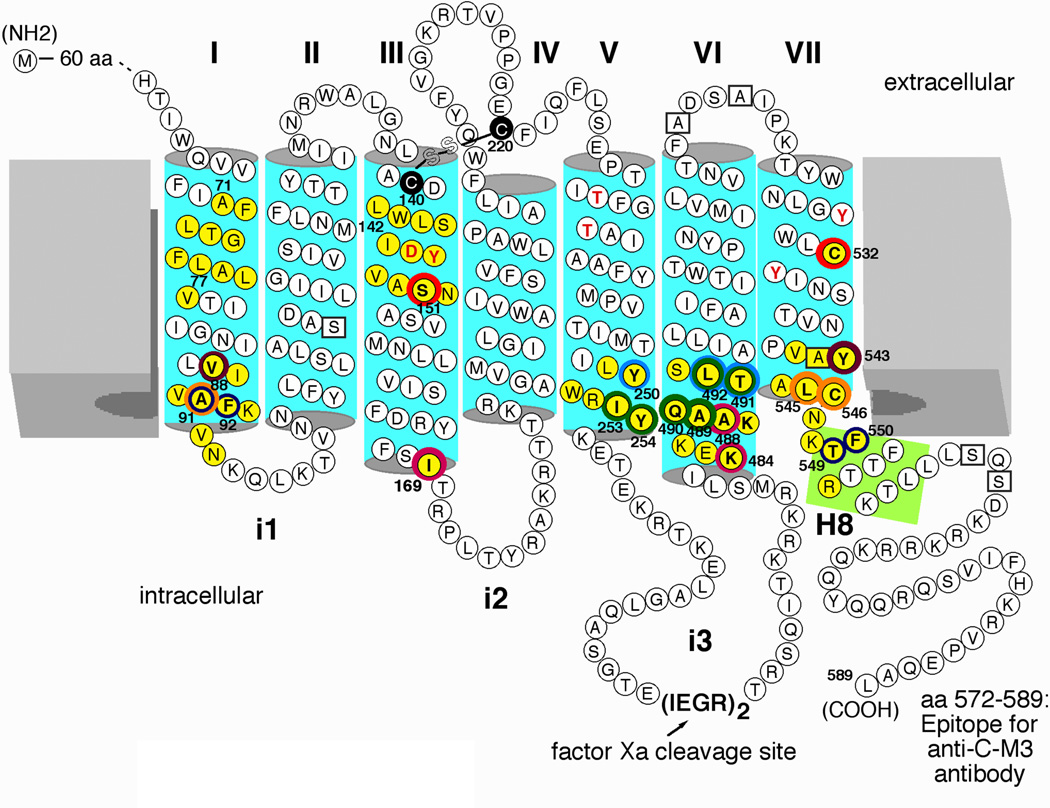
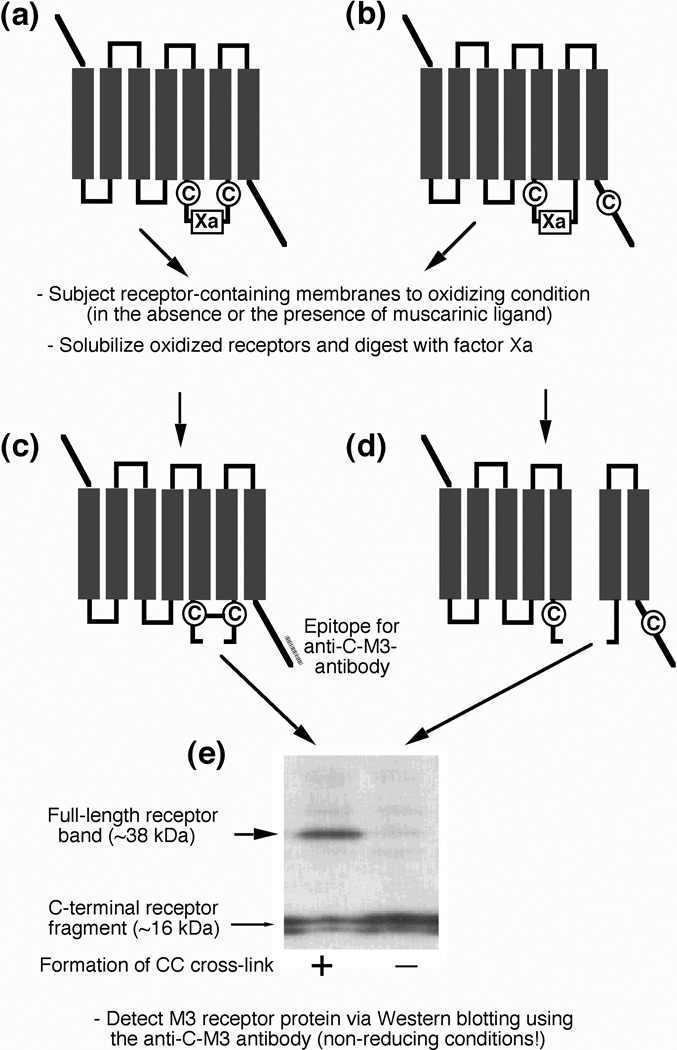
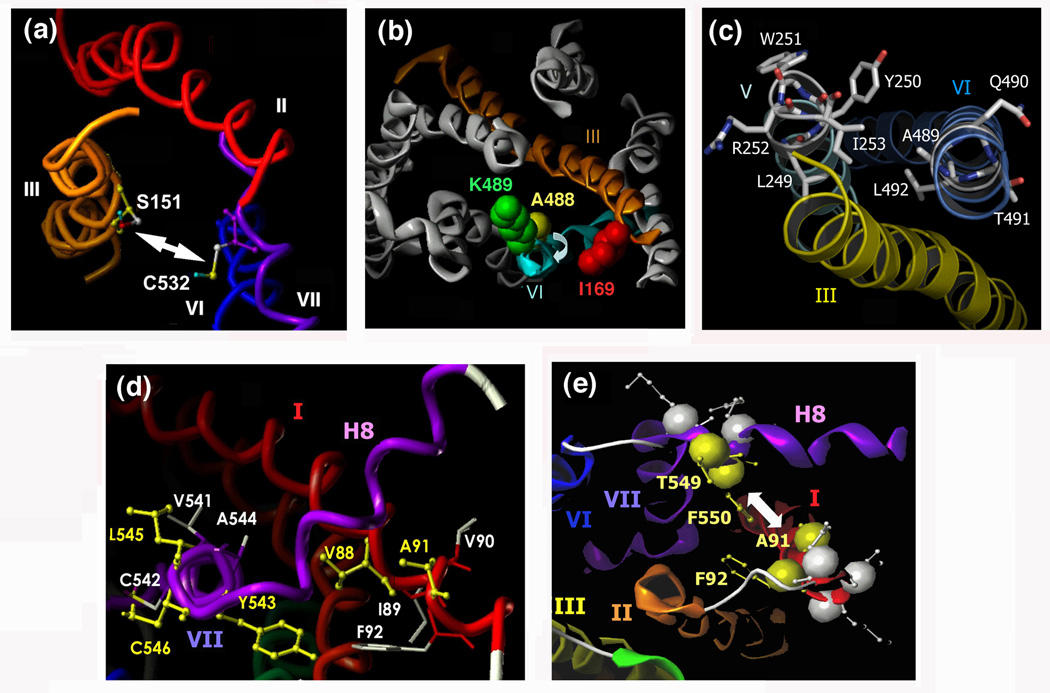
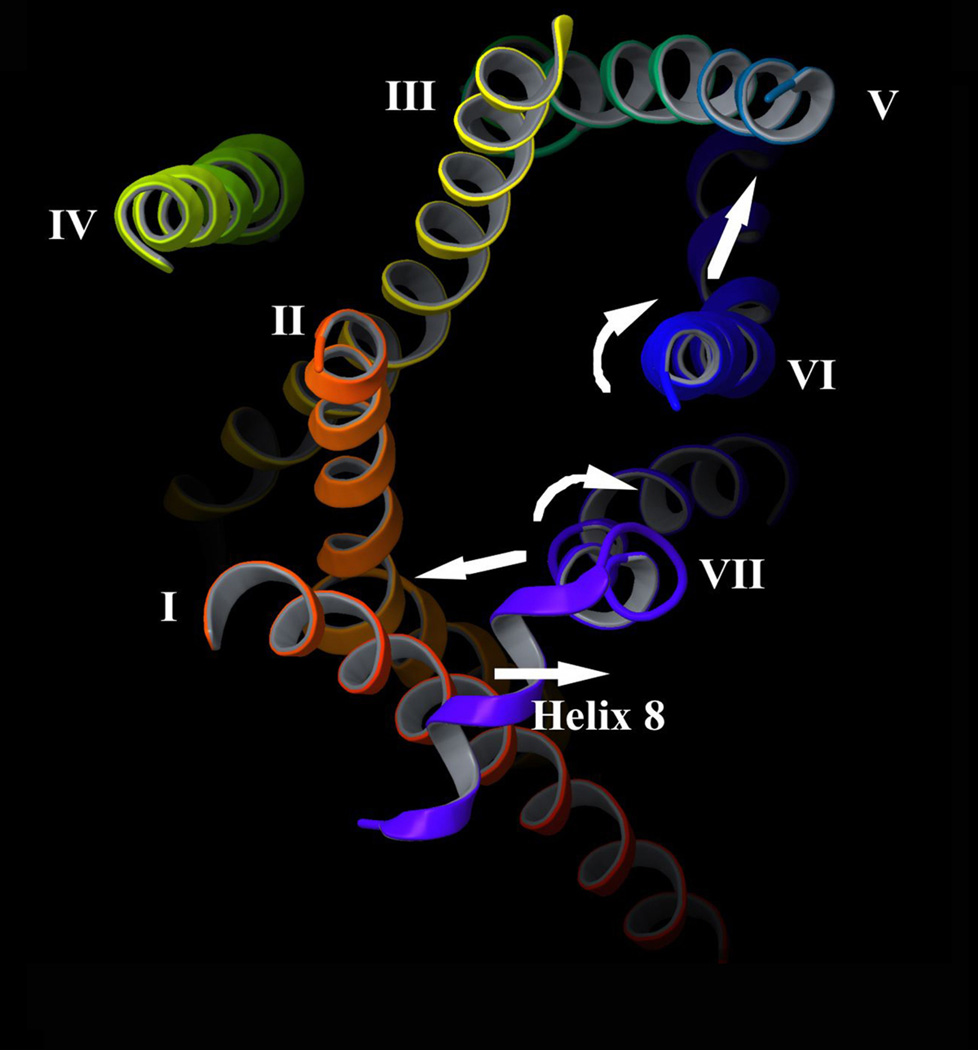
Little is known about the nature of the conformational changes that convert G-protein-coupled receptors (GPCRs), which bind diffusible ligands, from their resting into their active states. To gain structural insight into this process, various laboratories have used disulfide cross-linking strategies involving cysteine-substituted mutant GPCRs. Several recent disulfide cross-linking studies using the M(3) muscarinic acetylcholine receptor as a model system have led to novel insights into the conformational changes associated with the activation of this prototypical class I GPCR. These structural changes are predicted to involve multiple receptor regions, primarily distinct segments of transmembrane helices III, VI and VII and helix 8. Given the high degree of structural homology found among most GPCRs, it is likely that these findings will be of considerable general relevance. A better understanding of the molecular mechanisms underlying GPCR activation might lead to novel strategies aimed at modulating GPCR function for therapeutic purposes.
Figures
Similar articles
-
Ligand-specific changes in M3 muscarinic acetylcholine receptor structure detected by a disulfide scanning strategy.Biochemistry. 2008 Mar 4;47(9):2776-88. doi: 10.1021/bi7019113. Epub 2008 Feb 5. Biochemistry. 2008. PMID: 18247581
-
Use of an in situ disulfide cross-linking strategy to study the dynamic properties of the cytoplasmic end of transmembrane domain VI of the M3 muscarinic acetylcholine receptor.Biochemistry. 2006 Jan 24;45(3):676-85. doi: 10.1021/bi051503q. Biochemistry. 2006. PMID: 16411743
-
Pronounced conformational changes following agonist activation of the M(3) muscarinic acetylcholine receptor.J Biol Chem. 2005 Jul 1;280(26):24870-9. doi: 10.1074/jbc.M500379200. Epub 2005 May 2. J Biol Chem. 2005. PMID: 15870064
-
Novel insights into M3 muscarinic acetylcholine receptor physiology and structure.J Mol Neurosci. 2014 Jul;53(3):316-23. doi: 10.1007/s12031-013-0127-0. Epub 2013 Sep 26. J Mol Neurosci. 2014. PMID: 24068573 Free PMC article. Review.
-
Conformational changes in G-protein-coupled receptors-the quest for functionally selective conformations is open.Br J Pharmacol. 2008 Mar;153 Suppl 1(Suppl 1):S358-66. doi: 10.1038/sj.bjp.0707615. Epub 2007 Dec 3. Br J Pharmacol. 2008. PMID: 18059316 Free PMC article. Review.
Cited by
-
Homology modeling and molecular dynamics simulations of the active state of the nociceptin receptor reveal new insights into agonist binding and activation.Proteins. 2012 Aug;80(8):1948-61. doi: 10.1002/prot.24077. Epub 2012 May 17. Proteins. 2012. PMID: 22489047 Free PMC article.
-
Ligand-induced rearrangements of the GABA(B) receptor revealed by fluorescence resonance energy transfer.J Biol Chem. 2010 Apr 2;285(14):10291-9. doi: 10.1074/jbc.M109.077990. Epub 2010 Feb 3. J Biol Chem. 2010. PMID: 20129919 Free PMC article.
-
Arsenic requires sphingosine-1-phosphate type 1 receptors to induce angiogenic genes and endothelial cell remodeling.Am J Pathol. 2009 May;174(5):1949-58. doi: 10.2353/ajpath.2009.081016. Epub 2009 Apr 6. Am J Pathol. 2009. PMID: 19349368 Free PMC article.
-
Identification of disulfide bonds in protein proteolytic degradation products using de novo-protein unique sequence tags approach.J Proteome Res. 2010 Aug 6;9(8):4053-60. doi: 10.1021/pr1002559. J Proteome Res. 2010. PMID: 20590115 Free PMC article.
-
Exploring structural dynamics of a membrane protein by combining bioorthogonal chemistry and cysteine mutagenesis.Elife. 2019 Nov 12;8:e50776. doi: 10.7554/eLife.50776. Elife. 2019. PMID: 31714877 Free PMC article.
References
-
- Lagerström MC, Schiöth HB. Structural diversity of G protein-coupled receptors and significance for drug discovery. Nat. Rev. Drug Discov. 2008;7:339–357. - PubMed
-
- Palczewski K, et al. Crystal structure of rhodopsin: a G protein-coupled receptor. Science. 2000;289:739–745. - PubMed
-
- Rasmussen SG, et al. Crystal structure of the human β2 adrenergic G-proteincoupled receptor. Nature. 2007;450:383–387. - PubMed
-
- Rosenbaum DM, et al. GPCR engineering yields high-resolution structural insights into β2-adrenergic receptor function. Science. 2007;318:1266–1273. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
