

DcpS as a therapeutic target for spinal muscular atrophy
- PMID: 18839960
- PMCID: PMC2646632
- DOI: 10.1021/cb800120t
DcpS as a therapeutic target for spinal muscular atrophy
Abstract
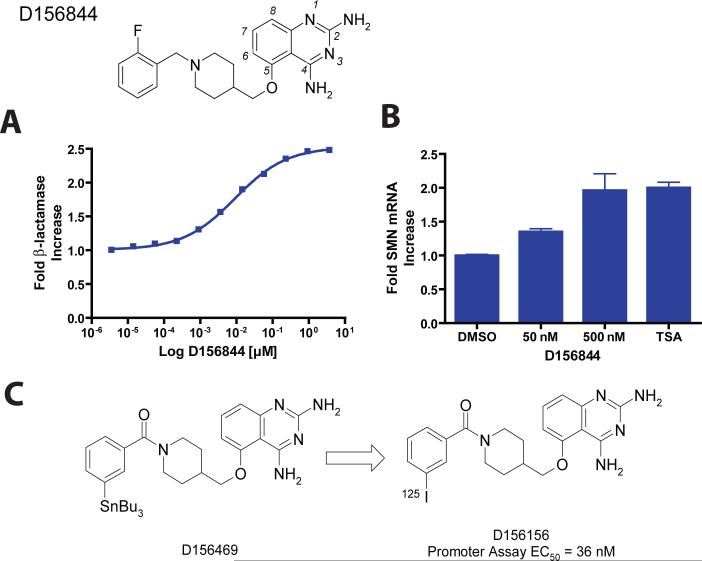
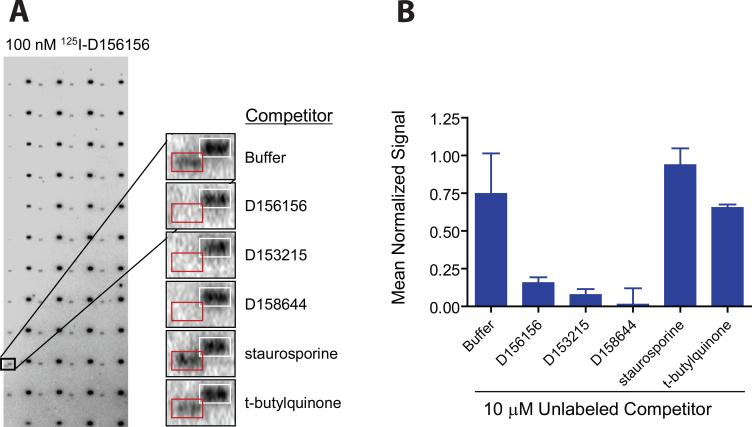
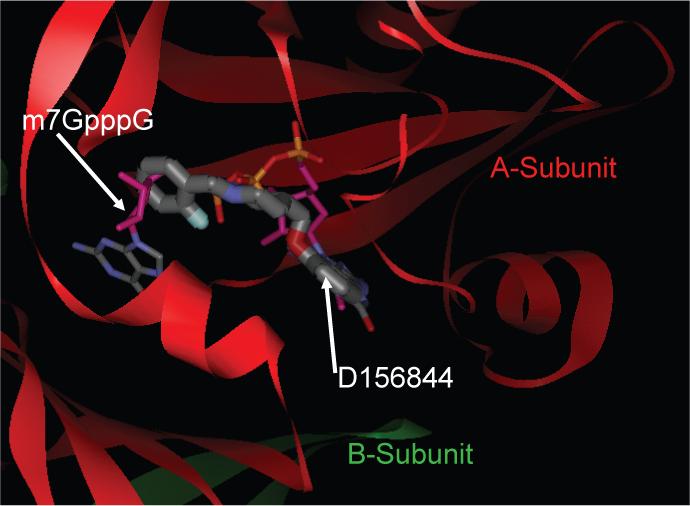
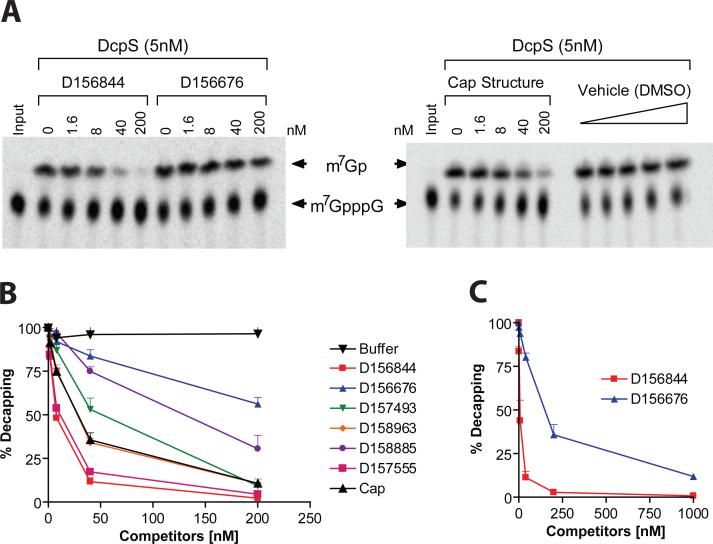
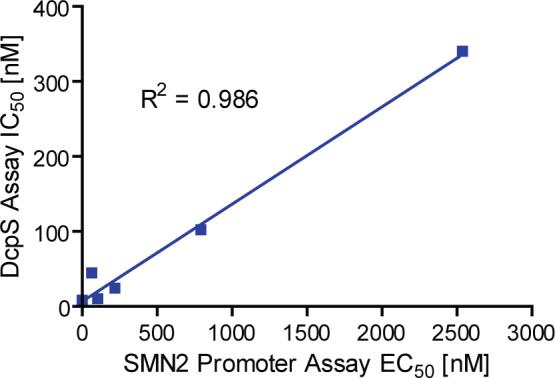
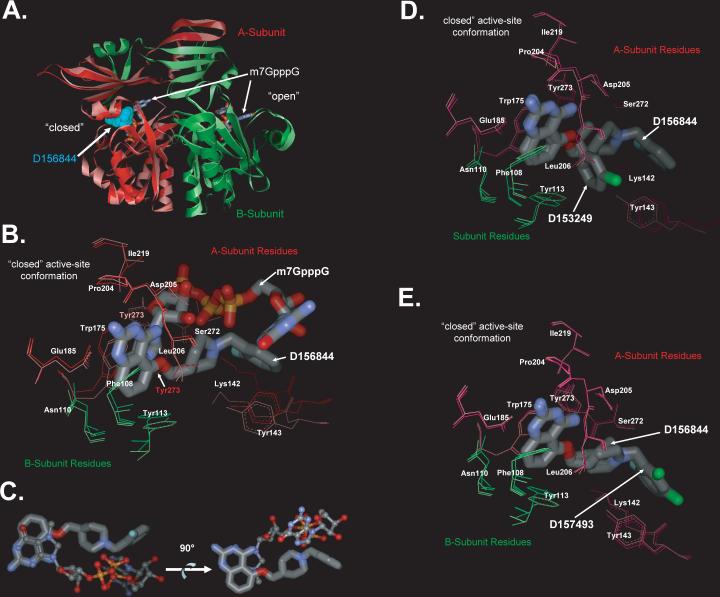
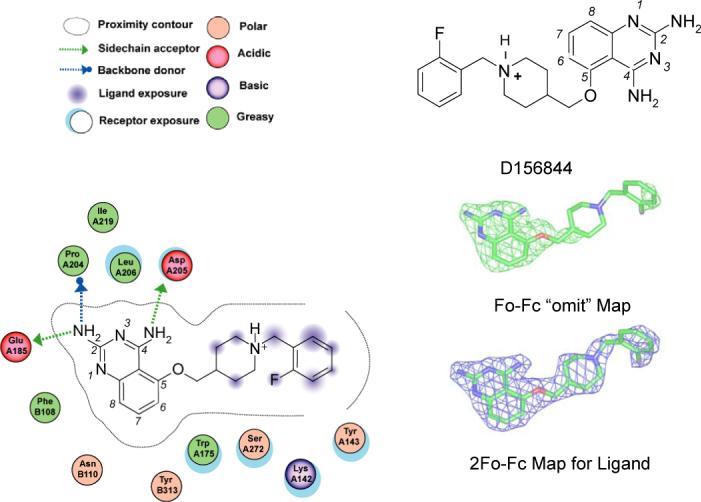
Spinal muscular atrophy (SMA) is caused by deletion or mutation of both copies of the SMN1 gene, which produces an essential protein known as SMN. The severity of SMA is modified by variable copy number of a second gene,SMN2, which produces an mRNA that is incorrectly spliced with deletion of the last exon. We described previously the discovery of potent C5-substituted quinazolines that increase SMN2 gene expression by 2-fold. Discovery of potent SMN2 promoter inducers relied on a cellular assay without knowledge of the molecular target. Using protein microarray scanning with a radiolabeled C5-substituted quinazoline probe, we identified the scavenger decapping enzyme, DcpS, as a potential binder. We show that the C5-substituted quinazolines potently inhibit DcpS decapping activity and that the potency of inhibition correlates with potency forSMN2 promoter induction. Binding of C5-substituted quinazolines to DcpS holds the enzyme in an open, catalytically incompetent conformation. DcpS is a nuclear shuttling protein that binds and hydrolyzes the m(7)GpppN mRNA cap structure and a modulator of RNA metabolism. Therefore DcpS represents a novel therapeutic target for modulating gene expression by a small molecule.
Figures
References
-
- Shiau AK, et al. The structural basis of estrogen receptor/coactivator recognition and the antagonism of this interaction by tamoxifen. Cell. 1998;95:927–37. - PubMed
-
- Kliewer SA, Lehmann JM, Milburn MV, Willson TM. The PPARs and PXRs: nuclear xenobiotic receptors that define novel hormone signaling pathways. Recent Prog Horm Res. 1999;54:345–67. discussion 367−8. - PubMed
-
- Lefebvre S, et al. Identification and characterization of a spinal muscular atrophy-determining gene. Cell. 1995;80:155–65. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Chemical Information
Medical
Molecular Biology Databases
Miscellaneous
