

Tubulin proteomics: towards breaking the code
- PMID: 18840397
- PMCID: PMC4039029
- DOI: 10.1016/j.ab.2008.09.020
Tubulin proteomics: towards breaking the code
Abstract
Since the discovery of tubulin as the major component of microtubules over 40 years ago, its diversity of forms has raised a continuum of fundamental questions about its regulation and functions in a variety of organisms across phyla. Its high abundance in the brain or in specialized organelles such as cilia has allowed early characterization of this important target for anticancer drugs. However, it was only when matrix-assisted laser desorption ionization and electrospray ionization mass spectrometry technologies became available in the late 1980's that the full complexity of tubulin expression patterns became more obvious. This contributed in a major way to the idea that due to increasing and conserved tubulin heterogeneity during evolution, a tubulin code read by microtubule associated proteins might exist and be of functional significance. We review here the merging of recent genetic and cell biology studies with proteomics to decipher this code and illustrate some of the tubulin proteomic approaches with new data generated in our laboratories.
Figures
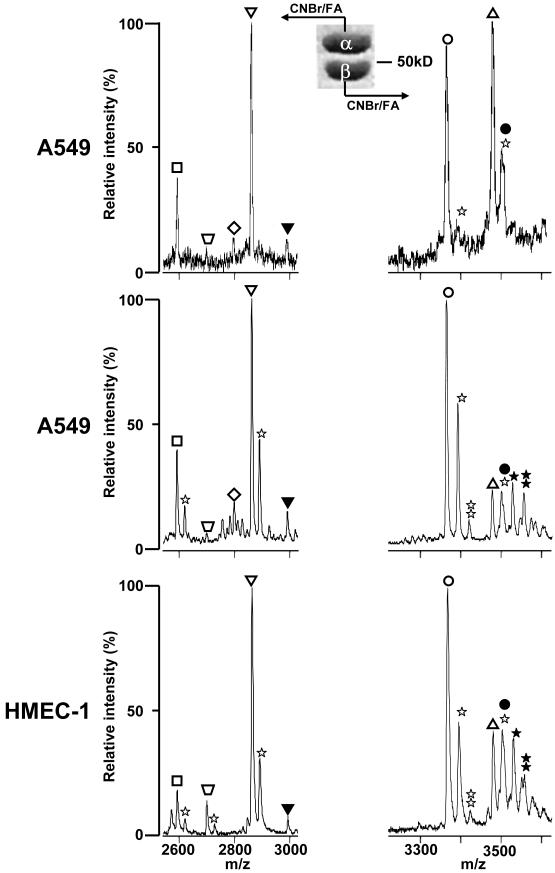
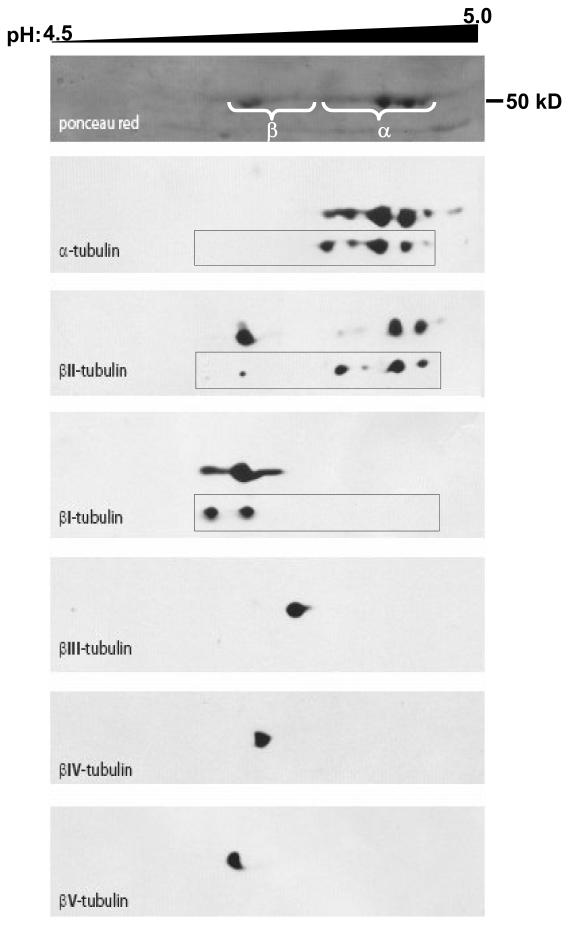
 , −163 Da), tyrosinated α4A (◇, +163 Da), α1B (∇), βI (◯), βIVb (△). The assignment of each m/z peak for β-tubulin is an interpretation based on the CNBr C-terminal peptides of βI- and βIVb-tubulin and their monoformylated derivatives.
, −163 Da), tyrosinated α4A (◇, +163 Da), α1B (∇), βI (◯), βIVb (△). The assignment of each m/z peak for β-tubulin is an interpretation based on the CNBr C-terminal peptides of βI- and βIVb-tubulin and their monoformylated derivatives.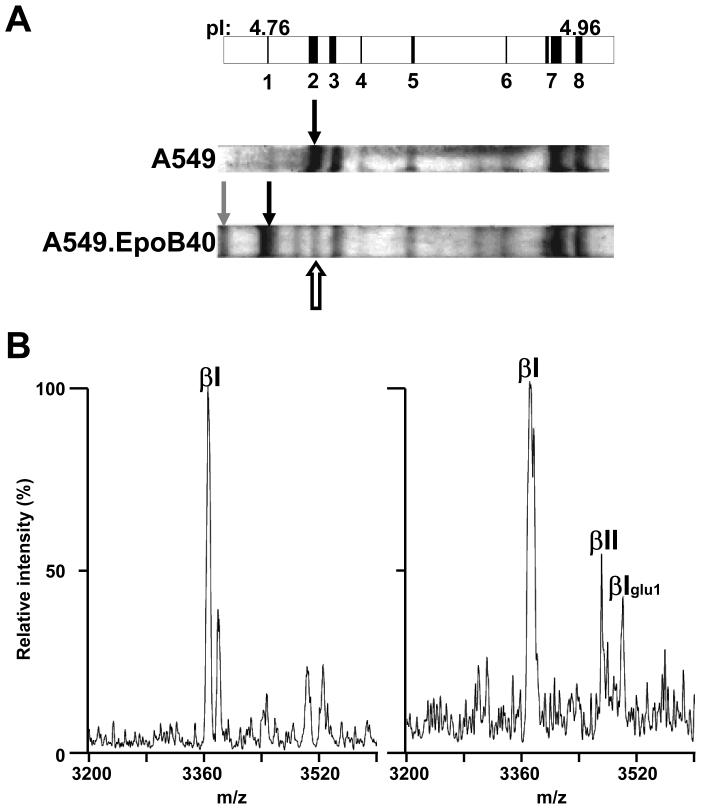
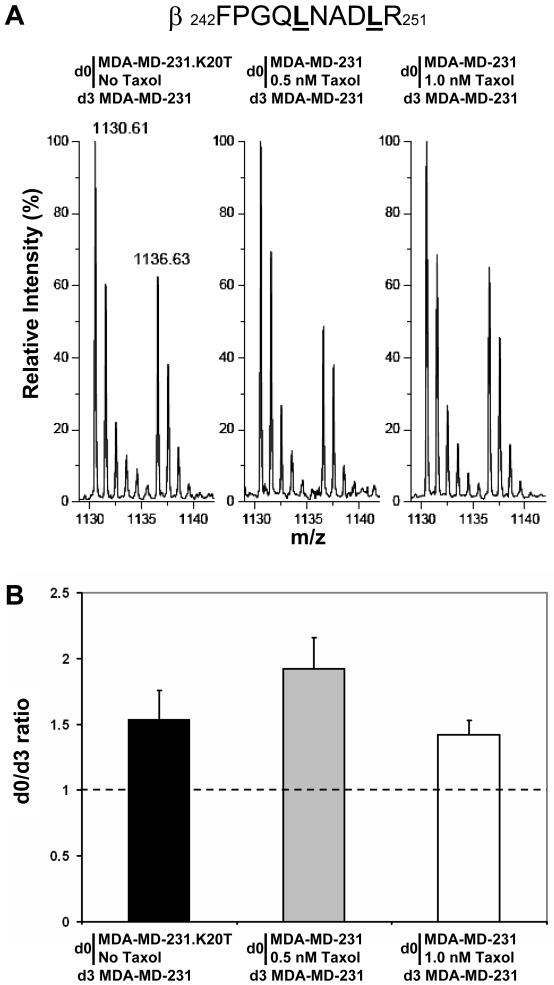
References
-
- Rodriguez OC, Schaefer AW, Mandato CA, Forscher P, Bement WM, Waterman-Storer CM. Conserved microtubule-actin interactions in cell movement and morphogenesis. Nat Cell Biol. 2003;5:599–609. - PubMed
-
- Bonnet C, Boucher D, Lazereg S, Pedrotti B, Islam K, Denoulet P, Larcher JC. Differential binding regulation of microtubule-associated proteins MAP1A, MAP1B, and MAP2 by tubulin polyglutamylation. J Biol Chem. 2001;276:12839–48. - PubMed
-
- Boucher D, Larcher JC, Gros F, Denoulet P. Polyglutamylation of tubulin as a progressive regulator of in vitro interactions between the microtubule-associated protein Tau and tubulin. Biochemistry. 1994;33:12471–7. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
