

The timing of differentiation of adult hippocampal neurons is crucial for spatial memory
- PMID: 18842068
- PMCID: PMC2561078
- DOI: 10.1371/journal.pbio.0060246
The timing of differentiation of adult hippocampal neurons is crucial for spatial memory
Abstract
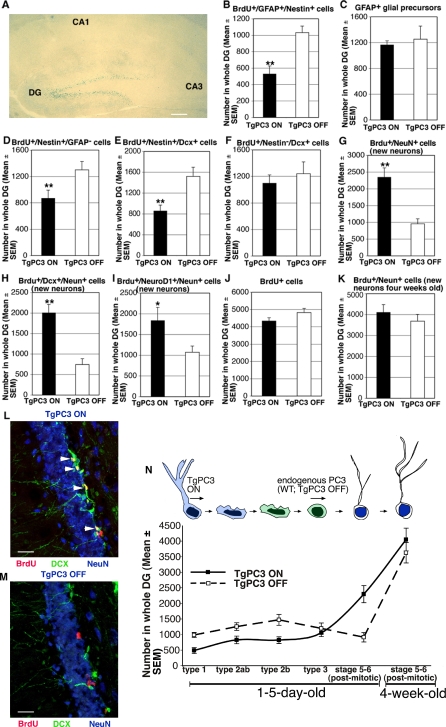
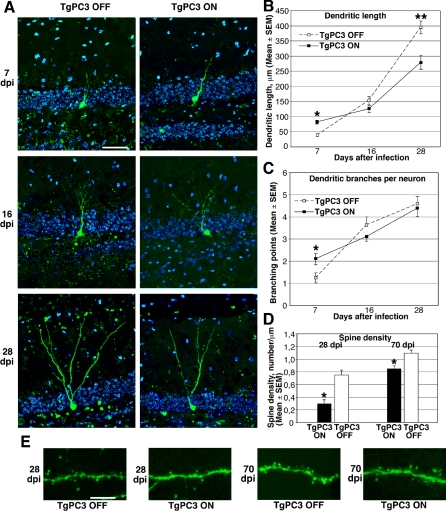
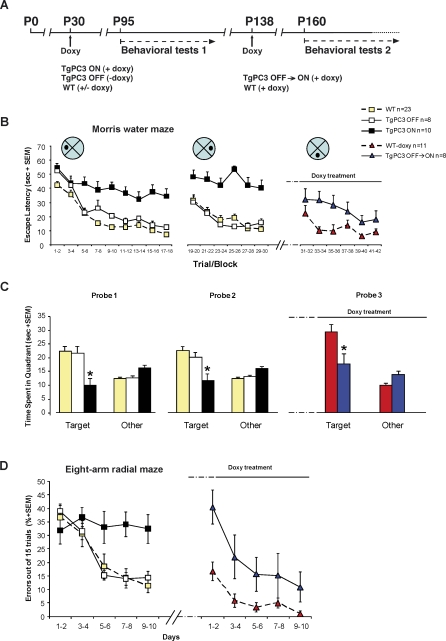
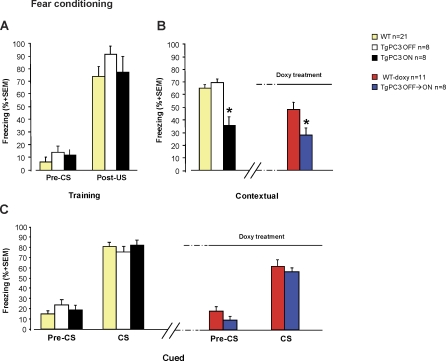
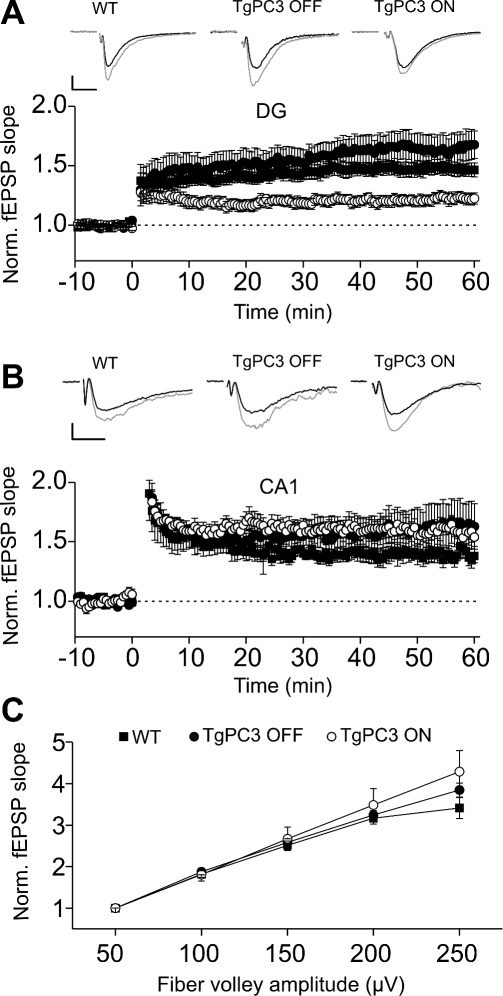
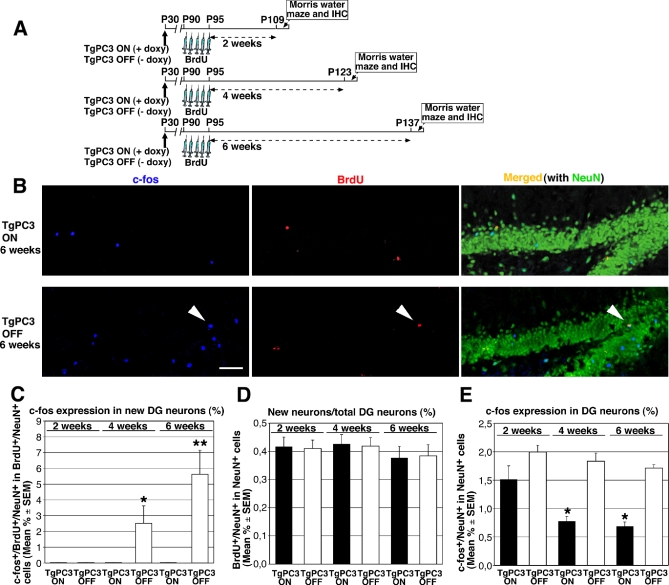
Adult neurogenesis in the dentate gyrus plays a critical role in hippocampus-dependent spatial learning. It remains unknown, however, how new neurons become functionally integrated into spatial circuits and contribute to hippocampus-mediated forms of learning and memory. To investigate these issues, we used a mouse model in which the differentiation of adult-generated dentate gyrus neurons can be anticipated by conditionally expressing the pro-differentiative gene PC3 (Tis21/BTG2) in nestin-positive progenitor cells. In contrast to previous studies that affected the number of newly generated neurons, this strategy selectively changes their timing of differentiation. New, adult-generated dentate gyrus progenitors, in which the PC3 transgene was expressed, showed accelerated differentiation and significantly reduced dendritic arborization and spine density. Functionally, this genetic manipulation specifically affected different hippocampus-dependent learning and memory tasks, including contextual fear conditioning, and selectively reduced synaptic plasticity in the dentate gyrus. Morphological and functional analyses of hippocampal neurons at different stages of differentiation, following transgene activation within defined time-windows, revealed that the new, adult-generated neurons up to 3-4 weeks of age are required not only to acquire new spatial information but also to use previously consolidated memories. Thus, the correct unwinding of these key memory functions, which can be an expression of the ability of adult-generated neurons to link subsequent events in memory circuits, is critically dependent on the correct timing of the initial stages of neuron maturation and connection to existing circuits.
Conflict of interest statement
Figures
Similar articles
-
Impaired terminal differentiation of hippocampal granule neurons and defective contextual memory in PC3/Tis21 knockout mice.PLoS One. 2009 Dec 17;4(12):e8339. doi: 10.1371/journal.pone.0008339. PLoS One. 2009. PMID: 20020054 Free PMC article.
-
Posttraining ablation of adult-generated neurons degrades previously acquired memories.J Neurosci. 2011 Oct 19;31(42):15113-27. doi: 10.1523/JNEUROSCI.3432-11.2011. J Neurosci. 2011. PMID: 22016545 Free PMC article.
-
Primary cilia regulate proliferation of amplifying progenitors in adult hippocampus: implications for learning and memory.J Neurosci. 2011 Jul 6;31(27):9933-44. doi: 10.1523/JNEUROSCI.1062-11.2011. J Neurosci. 2011. PMID: 21734285 Free PMC article.
-
Adult hippocampal neurogenesis, synaptic plasticity and memory: facts and hypotheses.Rev Neurosci. 2007;18(2):93-114. doi: 10.1515/revneuro.2007.18.2.93. Rev Neurosci. 2007. PMID: 17593874 Review.
-
The gene PC3(TIS21/BTG2), prototype member of the PC3/BTG/TOB family: regulator in control of cell growth, differentiation, and DNA repair?J Cell Physiol. 2001 May;187(2):155-65. doi: 10.1002/jcp.1062. J Cell Physiol. 2001. PMID: 11267995 Review.
Cited by
-
Hippocampal neurogenesis: Learning to remember.Prog Neurobiol. 2016 Mar-May;138-140:1-18. doi: 10.1016/j.pneurobio.2015.12.006. Epub 2016 Feb 6. Prog Neurobiol. 2016. PMID: 26855369 Free PMC article. Review.
-
Neural mechanisms underlying GABAergic regulation of adult hippocampal neurogenesis.Cell Tissue Res. 2018 Jan;371(1):33-46. doi: 10.1007/s00441-017-2668-y. Epub 2017 Sep 25. Cell Tissue Res. 2018. PMID: 28948349 Free PMC article. Review.
-
Neurogenic Effects of Low-Dose Whole-Body HZE (Fe) Ion and Gamma Irradiation.Radiat Res. 2016 Dec;186(6):614-623. doi: 10.1667/RR14530.1. Epub 2016 Dec 1. Radiat Res. 2016. PMID: 27905869 Free PMC article.
-
Brain-derived neurotrophic factor signaling in the HVC is required for testosterone-induced song of female canaries.J Neurosci. 2009 Dec 9;29(49):15511-9. doi: 10.1523/JNEUROSCI.2564-09.2009. J Neurosci. 2009. PMID: 20007475 Free PMC article.
-
Basolateral amygdala regulation of adult hippocampal neurogenesis and fear-related activation of newborn neurons.Mol Psychiatry. 2012 May;17(5):527-36. doi: 10.1038/mp.2011.71. Epub 2011 Jun 14. Mol Psychiatry. 2012. PMID: 21670733 Free PMC article.
References
-
- Altman J, Das GD. Autoradiographic and histological evidence of post-natal hippocampal neurogenesis in rats. J Comp Neurol. 1965;124:319–335. - PubMed
-
- Nottebohm F. Neuronal replacement in adult brain. Brain Res Bull. 2002;57:737–749. - PubMed
-
- Bayer SA, Yackel JW, Puri PS. Neurons in the rat dentate gyrus granular layer substantially increase during juvenile and adult life. Science. 1982;216:890–892. - PubMed
-
- Eriksson PS, Perfilieva E, Bjork-Eriksson T, Alborn AM, Nordborg C, et al. Neurogenesis in the adult human hippocampus. Nat Med. 1998;4:1313–1317. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
