

APOBEC3G subunits self-associate via the C-terminal deaminase domain
- PMID: 18842592
- PMCID: PMC2586250
- DOI: 10.1074/jbc.M803726200
APOBEC3G subunits self-associate via the C-terminal deaminase domain
Abstract
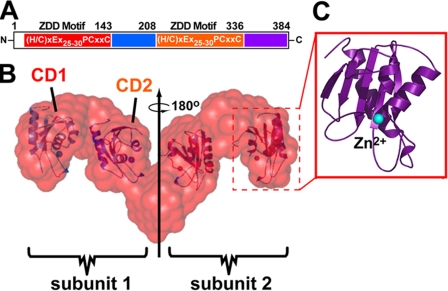
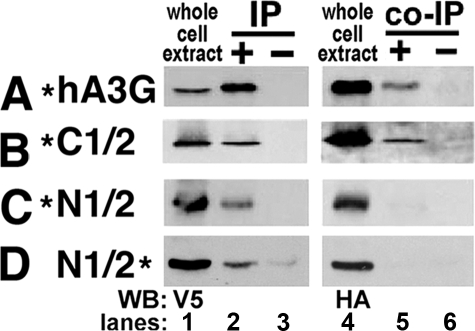
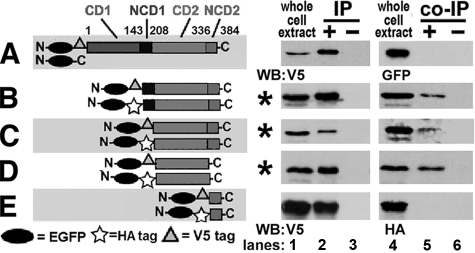
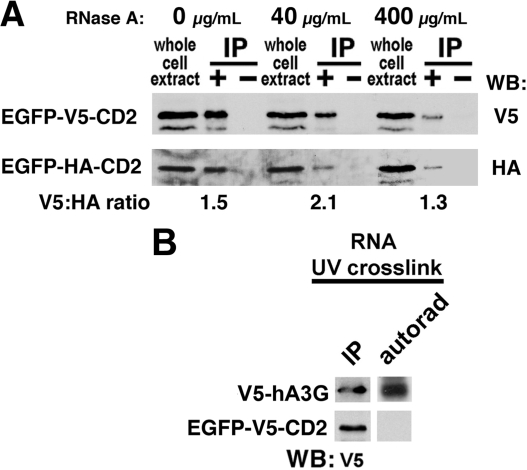
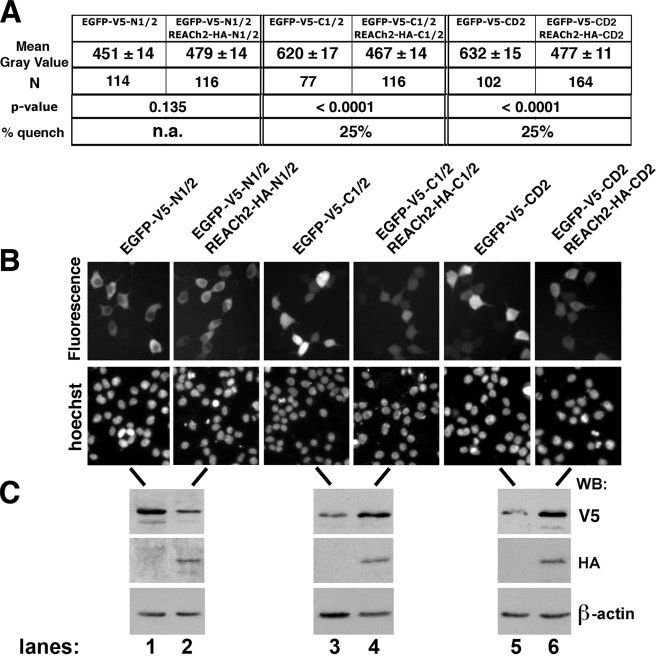
Human APOBEC3G (hA3G) is a cytidine deaminase active on HIV single-stranded DNA. Small angle x-ray scattering and molecular envelope restorations predicted a C-terminal dimeric model for RNA-depleted hA3G in solution. Each subunit was elongated, suggesting that individual domains of hA3G are solvent-exposed and therefore may interact with other macromolecules even as isolated substructures. In this study, co-immunoprecipitation and in-cell quenched fluorescence resonance energy transfer assays reveal that hA3G forms RNA-independent oligomers through interactions within its C terminus. Residues 209-336 were necessary and sufficient for homoligomerization. N-terminal domains of hA3G were unable to multimerize but remained functional for Gag and viral infectivity factor (Vif) interactions when expressed apart from the C terminus. These findings corroborate the small angle x-ray scattering structural model and are instructive for development of high throughput screens that target specific domains and their functions to identify HIV/AIDS therapeutics.
Figures
References
-
- MacElrevey, C., and Wedekind, J. E. (2008) in RNA and DNA Editing: Molecular Mechanisms and Their Integration into Biological Systems (Smith, H. C., ed) pp. 369-419, Wiley, New York
-
- Wedekind, J. E., Dance, G. S., Sowden, M. P., and Smith, H. C. (2003) Trends Genet. 19 207-216 - PubMed
-
- Jarmuz, A., Chester, A., Bayliss, J., Gisbourne, J., Dunham, I., Scott, J., and Navaratnam, N. (2002) Genomics 79 285-296 - PubMed
-
- Sheehy, A., Gaddis, N., Choi, J., and Malim, M. (2002) Nature 418 646-650 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
