

The p300 acetylase is critical for ligand-activated farnesoid X receptor (FXR) induction of SHP
- PMID: 18842595
- PMCID: PMC2596376
- DOI: 10.1074/jbc.M803531200
The p300 acetylase is critical for ligand-activated farnesoid X receptor (FXR) induction of SHP
Abstract
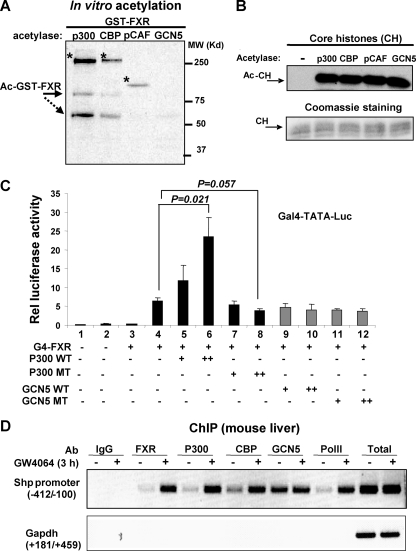
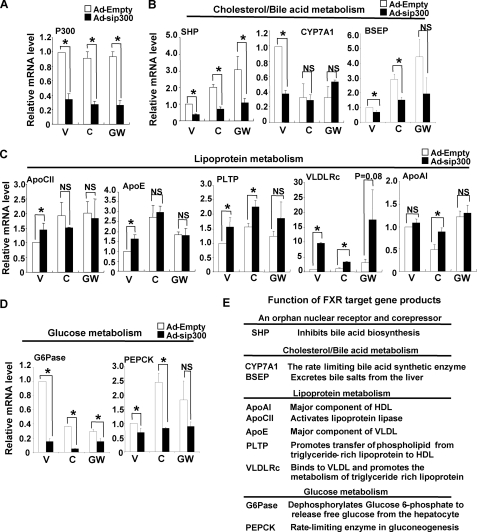
The primary bile acid receptor farnesoid X receptor (FXR) maintains lipid and glucose homeostasis by regulating expression of numerous bile acid-responsive genes, including an orphan nuclear receptor and metabolic regulator SHP. Using SHP as a model gene, we studied how FXR activity is regulated by p300 acetylase. FXR interaction with p300 and their recruitment to the SHP promoter and acetylated histone levels at the promoter were increased by FXR agonists in mouse liver and HepG2 cells. In contrast, p300 recruitment and acetylated histones at the promoter were not detected in FXR-null mice. p300 directly interacted with and acetylated FXR in vitro. Overexpression of p300 wild type increased, whereas a catalytically inactive p300 mutant decreased, acetylated FXR levels and FXR transactivation in cells. While similar results were observed with a related acetylase, CBP, GCN5 did not enhance FXR transactivation, and its recruitment to the promoter was not increased by FXR agonists, suggesting functional specificity of acetylases in FXR signaling. Down-regulation of p300 by siRNA decreased acetylated FXR and acetylated histone levels, and occupancy of FXR at the promoter, resulting in substantial inhibition of SHP expression. These results indicate that p300 acts as a critical coactivator of FXR induction of SHP by acetylating histones at the promoter and FXR itself. Surprisingly, p300 down-regulation altered expression of other metabolic FXR target genes involved in lipoprotein and glucose metabolism, such that beneficial lipid and glucose profiles would be expected. These unexpected findings suggest that inhibition of hepatic p300 activity may be beneficial for treating metabolic diseases.
Figures







References
-
- Makishima, M., Okamoto, A. Y., Repa, J. J., Tu, H., Learned, M., Luk, A., Hull, M. V., Lustig, K. D., Mangelsdorf, D. J., and Shan, B. (1999) Science 284 1362-1365 - PubMed
-
- Mangelsdorf, D. J., and Evans, R. M. (1995) Cell 83 841-850 - PubMed
-
- Wang, H., Chen, J., Hollister, K., Sowers, L., and Forman, B. M. (1999) Mol. Cell 3 543-553 - PubMed
-
- Parks, D. J., Blanchard, S. G., Bledsoe, R. K., Chandra, G., Consler, T. G., Kliewer, S. A., Stimmel, J. B., Willson, T. M., Zavacki, A. M., Moore, D. D., and Lehmann, J. M. (1999) Science 284 1365-1368 - PubMed
-
- Kalaany, N. Y., and Mangelsdorf, D. J. (2006) Annu. Rev. Physiol. 68 159-191 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
