

BDNF modulation of NMDA receptors is activity dependent
- PMID: 18842955
- PMCID: PMC2604850
- DOI: 10.1152/jn.90418.2008
BDNF modulation of NMDA receptors is activity dependent
Abstract
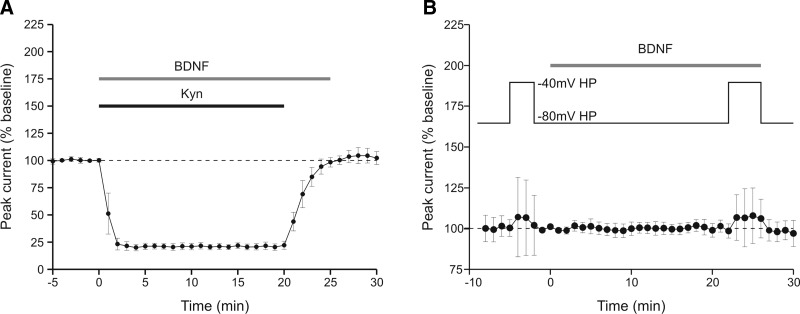
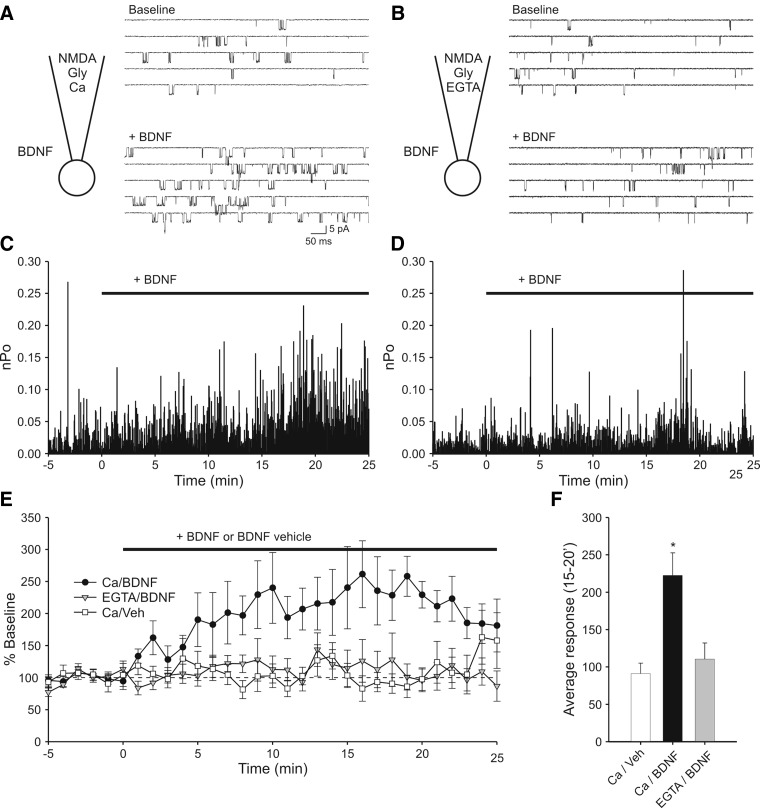
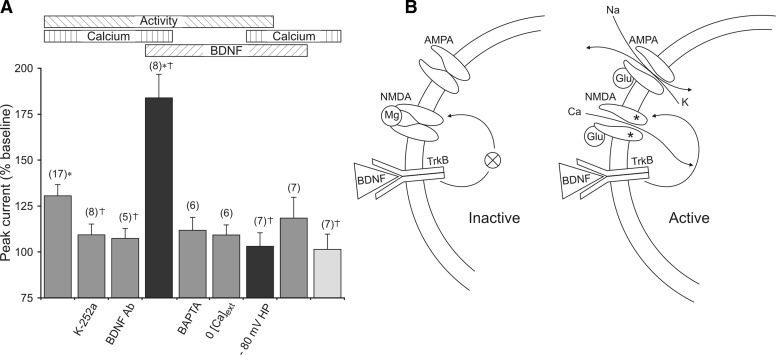
Brain-derived neurotrophic factor (BDNF), a potent modulator of synaptic transmission, is known to influence associative synaptic plasticity and refinement of neural connectivity. We now show that BDNF modulation of glutamate currents in hippocampal neurons exhibits the additional property of use dependence, a postsynaptic mechanism resulting in selective modulation of active channels. We demonstrate selectivity by varying the repetition rate of iontophoretically applied glutamate pulses during BDNF exposure. During relatively high-frequency glutamate pulses (0.1 Hz), BDNF application elicited a doubling of the glutamate current. During low-frequency pulses (0.0033 Hz), however, BDNF evoked a dramatically diminished response. This effect was apparently mediated by calcium because manipulations that prevented elevation of intracellular calcium largely eliminated the action of BDNF on glutamate currents. To confirm N-methyl-D-aspartate (NMDA) receptor involvement and assess spatial requirements, we made cell-attached single-channel recordings from somatic NMDA receptors. Inclusion of calcium in the pipette was sufficient to produce enhancement of channel activity by BDNF. Substitution of EGTA for calcium prevented BDNF effects. We conclude that BDNF modulation of postsynaptic NMDA receptors requires concurrent neuronal activity potentially conferring synaptic specificity on the neurotrophin's actions.
Figures
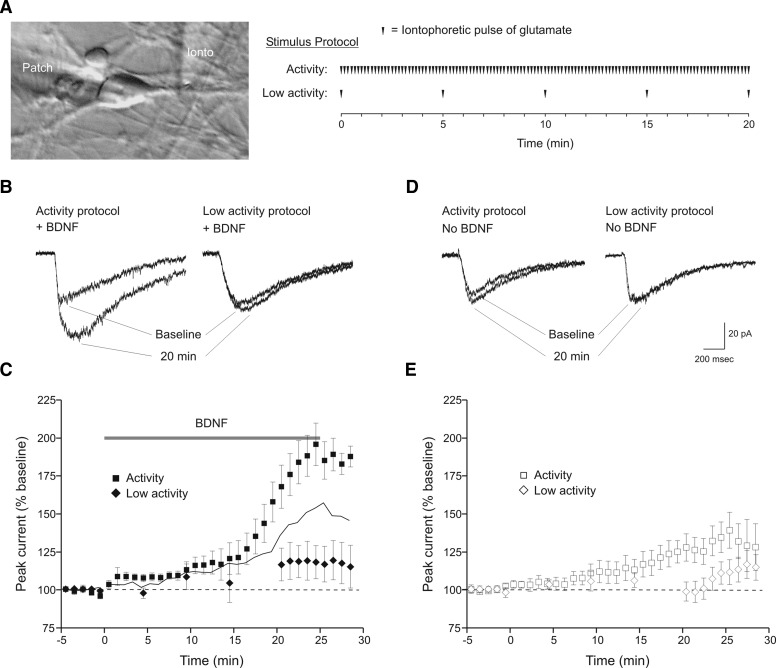
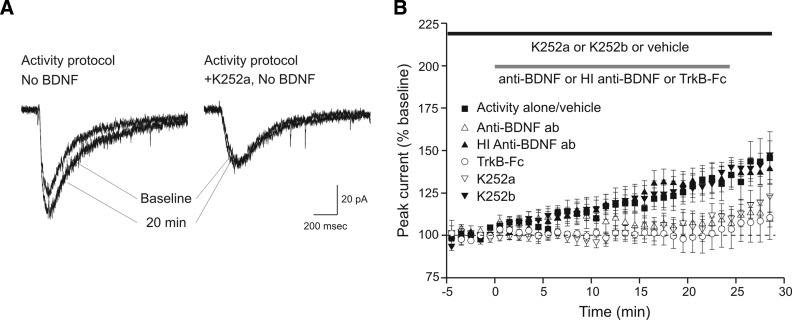
 and □ (bars), application of the indicated compounds in separate sets of experiments. For all conditions, the activity protocol (0.1-Hz stimulation frequency) was used.
and □ (bars), application of the indicated compounds in separate sets of experiments. For all conditions, the activity protocol (0.1-Hz stimulation frequency) was used.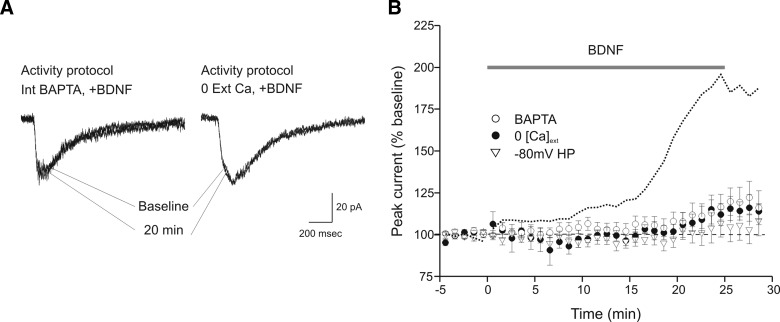
Similar articles
-
Neurotrophin modulation of NMDA receptors in cultured murine and isolated rat neurons.J Neurophysiol. 1997 Nov;78(5):2363-71. doi: 10.1152/jn.1997.78.5.2363. J Neurophysiol. 1997. PMID: 9356388
-
Blockade of NR2B-containing NMDA receptors prevents BDNF enhancement of glutamatergic transmission in hippocampal neurons.Learn Mem. 1999 May-Jun;6(3):257-66. Learn Mem. 1999. PMID: 10492007 Free PMC article.
-
The excitoprotective effect of N-methyl-D-aspartate receptors is mediated by a brain-derived neurotrophic factor autocrine loop in cultured hippocampal neurons.J Neurochem. 2005 Aug;94(3):713-22. doi: 10.1111/j.1471-4159.2005.03200.x. Epub 2005 Jul 5. J Neurochem. 2005. PMID: 16000165
-
Presynaptic and postsynaptic NMDA receptors mediate distinct effects of brain-derived neurotrophic factor on synaptic transmission.J Neurophysiol. 2008 Dec;100(6):3175-84. doi: 10.1152/jn.90880.2008. Epub 2008 Oct 15. J Neurophysiol. 2008. PMID: 18922945 Free PMC article.
-
Modulation of the NMDA Receptor Through Secreted Soluble Factors.Mol Neurobiol. 2016 Jan;53(1):299-309. doi: 10.1007/s12035-014-9009-x. Epub 2014 Nov 29. Mol Neurobiol. 2016. PMID: 25429903 Review.
Cited by
-
Acute intermittent hypoxia boosts spinal plasticity in humans with tetraplegia.Exp Neurol. 2021 Jan;335:113483. doi: 10.1016/j.expneurol.2020.113483. Epub 2020 Sep 25. Exp Neurol. 2021. PMID: 32987000 Free PMC article.
-
BDNF Contributes to Spinal Long-Term Potentiation and Mechanical Hypersensitivity Via Fyn-Mediated Phosphorylation of NMDA Receptor GluN2B Subunit at Tyrosine 1472 in Rats Following Spinal Nerve Ligation.Neurochem Res. 2017 Oct;42(10):2712-2729. doi: 10.1007/s11064-017-2274-0. Epub 2017 May 11. Neurochem Res. 2017. PMID: 28497343
-
Synaptic regulation of affective behaviors; role of BDNF.Neuropharmacology. 2014 Jan;76 Pt C(0 0):684-95. doi: 10.1016/j.neuropharm.2013.04.011. Epub 2013 Jun 6. Neuropharmacology. 2014. PMID: 23747574 Free PMC article. Review.
-
Enriched environment attenuates changes in water-maze performance and BDNF level caused by prenatal alcohol exposure.EXCLI J. 2014 May 15;13:536-47. eCollection 2014. EXCLI J. 2014. PMID: 26417281 Free PMC article.
-
Neuron-Derived Extracellular Vesicles miRNA Profiles Identify Children Who Experience Adverse Events after Ketamine Administration for Procedural Sedation.Clin Pharmacol Ther. 2025 Jan;117(1):174-183. doi: 10.1002/cpt.3420. Epub 2024 Aug 20. Clin Pharmacol Ther. 2025. PMID: 39164873 Free PMC article.
References
-
- Arvanian VL, Mendell LM. Acute modulation of synaptic transmission to motoneurons by BDNF in the neonatal rat spinal cord. Eur J Neurosci 14: 1800–1808, 2001. - PubMed
-
- Arvanov VL, Seebach BS, Mendell LM. NT-3 Evokes an LTP-like facilitation of AMPA/Kainate receptor-mediated synaptic transmission in the neonatal rat spinal cord. J Neurophysiol 84: 752–758, 2000. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources