

Beclin 1 forms two distinct phosphatidylinositol 3-kinase complexes with mammalian Atg14 and UVRAG
- PMID: 18843052
- PMCID: PMC2592660
- DOI: 10.1091/mbc.e08-01-0080
Beclin 1 forms two distinct phosphatidylinositol 3-kinase complexes with mammalian Atg14 and UVRAG
Abstract
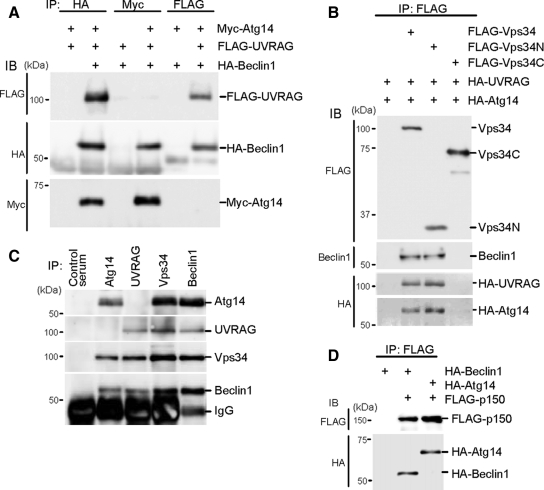
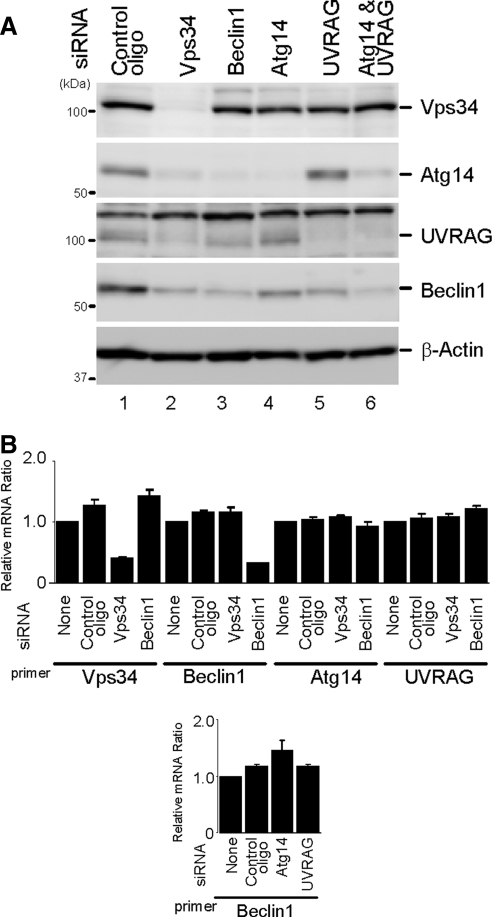
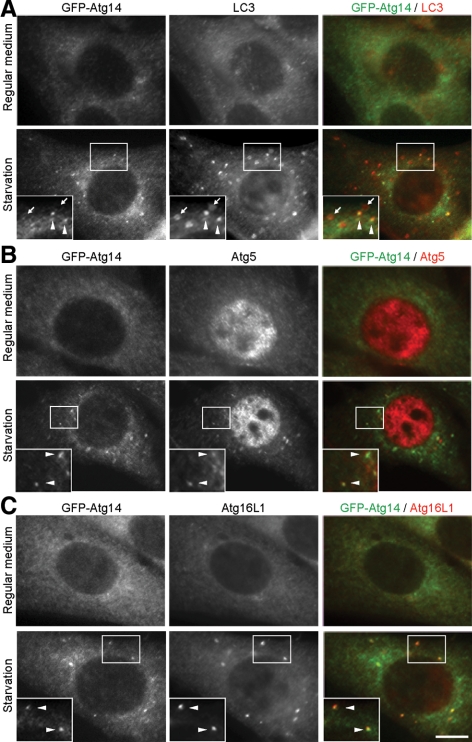
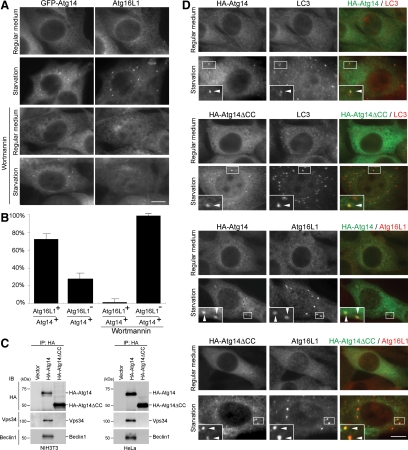
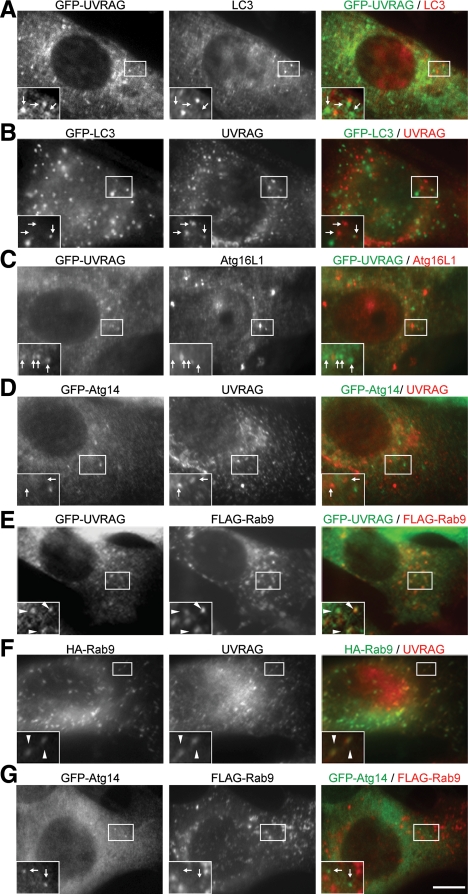
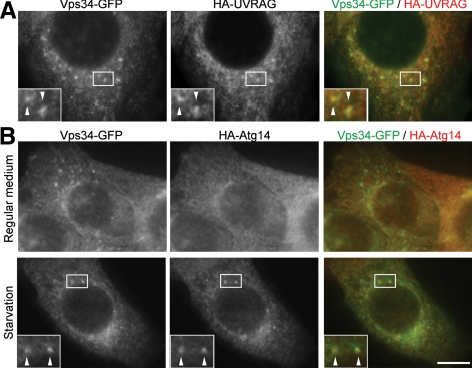
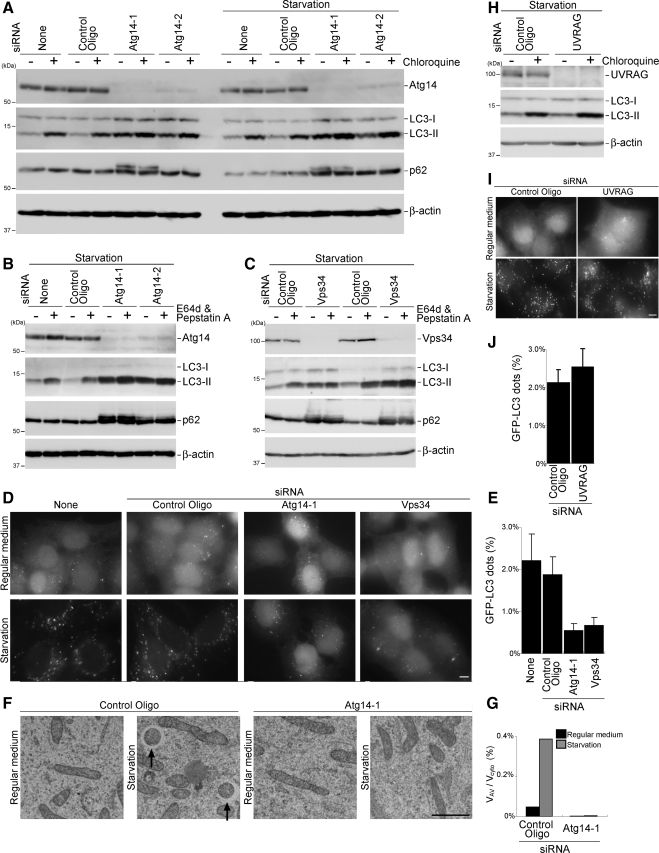
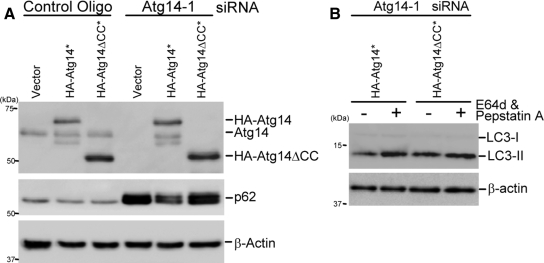
Class III phosphatidylinositol 3-kinase (PI3-kinase) regulates multiple membrane trafficking. In yeast, two distinct PI3-kinase complexes are known: complex I (Vps34, Vps15, Vps30/Atg6, and Atg14) is involved in autophagy, and complex II (Vps34, Vps15, Vps30/Atg6, and Vps38) functions in the vacuolar protein sorting pathway. Atg14 and Vps38 are important in inducing both complexes to exert distinct functions. In mammals, the counterparts of Vps34, Vps15, and Vps30/Atg6 have been identified as Vps34, p150, and Beclin 1, respectively. However, orthologues of Atg14 and Vps38 remain unknown. We identified putative mammalian homologues of Atg14 and Vps38. The Vps38 candidate is identical to UV irradiation resistance-associated gene (UVRAG), which has been reported as a Beclin 1-interacting protein. Although both human Atg14 and UVRAG interact with Beclin 1 and Vps34, Atg14, and UVRAG are not present in the same complex. Although Atg14 is present on autophagic isolation membranes, UVRAG primarily associates with Rab9-positive endosomes. Silencing of human Atg14 in HeLa cells suppresses autophagosome formation. The coiled-coil region of Atg14 required for binding with Vps34 and Beclin 1 is essential for autophagy. These results suggest that mammalian cells have at least two distinct class III PI3-kinase complexes, which may function in different membrane trafficking pathways.
Figures

References
-
- Berg T. O., Fengsrud M., Stromhaug P. E., Berg T., Seglen P. O. Isolation and characterization of rat liver amphisomes. Evidence for fusion of autophagosomes with both early and late endosomes. J. Biol. Chem. 1998;273:21883–21892. - PubMed
-
- Blommaart E. F., Krause U., Schellens J. P., Vreeling-Sindelarova H., Meijer A. J. The phosphatidylinositol 3-kinase inhibitors wortmannin and LY294002 inhibit autophagy in isolated rat hepatocytes. Eur. J. Biochem. 1997;243:240–246. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
