

Review
doi: 10.1038/nri2417.
Mammalian glycosylation in immunity
Affiliations
- PMID: 18846099
- PMCID: PMC2768770
- DOI: 10.1038/nri2417
Item in Clipboard
Review
Mammalian glycosylation in immunity
Nat Rev Immunol.
2008 Nov.
Abstract
Glycosylation produces a diverse and abundant repertoire of glycans, which are collectively known as the glycome. Glycans are one of the four fundamental macromolecular components of all cells, and are highly regulated in the immune system. Their diversity reflects their multiple biological functions that encompass ligands for proteinaceous receptors known as lectins. Since the discovery that selectins and their glycan ligands are important for the regulation of leukocyte trafficking, it has been shown that additional features of the vertebrate immune system are also controlled by endogenous cellular glycosylation. This Review focuses on the emerging immunological roles of the mammalian glycome.
Figures
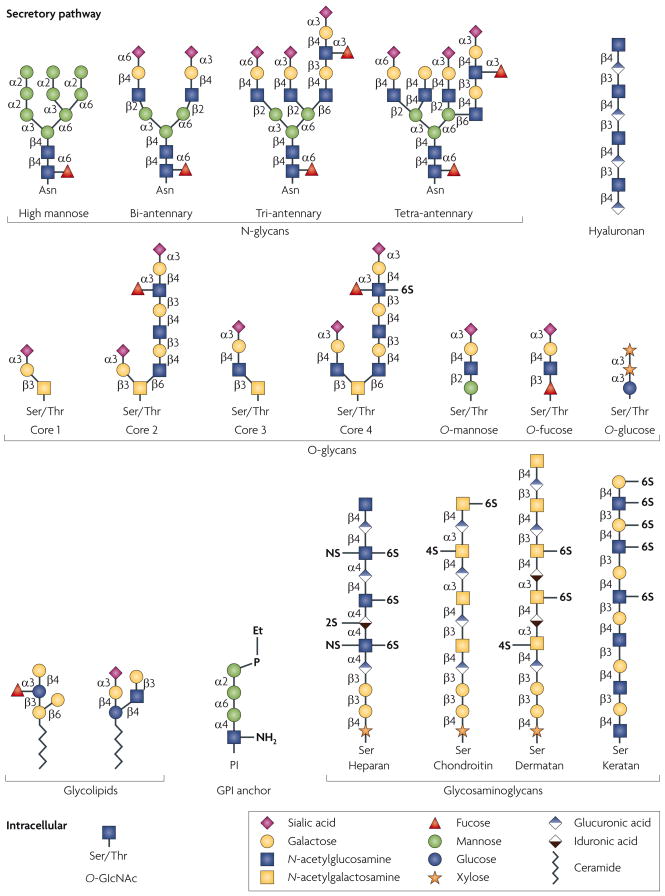
Glycan structures of the six classes of secretory glycan (N-glycans, hyaluronan, O-glycans, glycolipids, glycosylphosphatidylinositol (GPI) anchor and glycosaminoglycans) and the single intracellular glycan O-linked β-N-acetylglucosamine (O-GlcNAc) are shown. Representative examples of each type are indicated using the symbol nomenclature for monosaccharides (see key). Multiple and multi-antennary examples of the predominant mature, complex-type N-glycans are shown, including the high-mannose N-glycan that is found attached to some glycoproteins. Examples of core 1–4 O-glycans are depicted, as well as O-mannose, O-fucose and O-glucose structures. Core 5–7 O-glycans are not shown. The GPI anchor and examples of the glycosaminoglycans and glycolipids are also depicted. In all examples, glycan linkages are identified by the anomeric configuration (α orβ) of the donor saccharide following its linkage to the ring position (1–6) of the glycan acceptor. Et–P denotes a phosphoethanolamine linkage. NS, 2S, 4S and 6S denote the sulphation positions of the glycosaminoglycan chains. Asn, asparagine; Ser, serine; Thr, threonine.
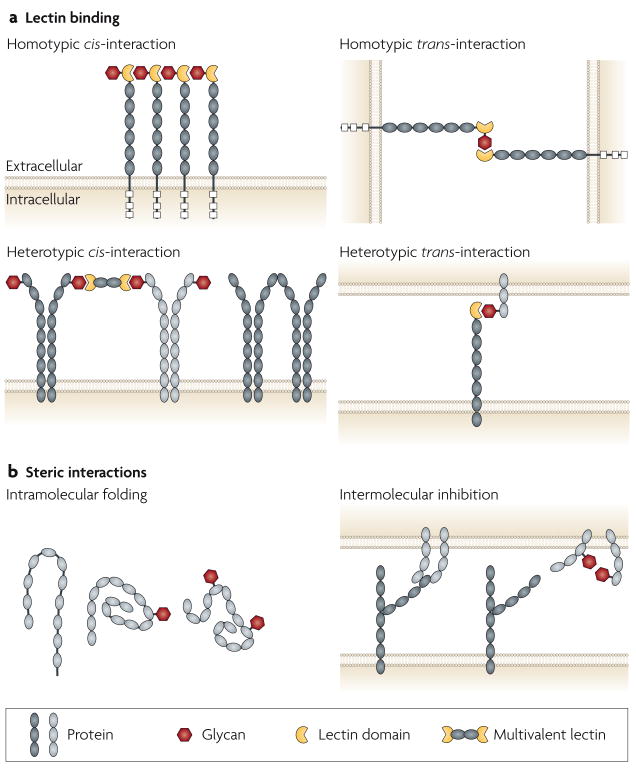
Glycans can function as ligands for lectins (a) and as steric elements that alter molecular interactions at the cell surface and between extracellular compartments (b). Interactions can involve the same cell (cis) or different cells (trans), and can be either homotypic or heterotypic, and intramolecular or intermolecular. It is often difficult to distinguish steric effects of glycans from alterations in lectin–glycan binding. This figure is idealized and not drawn to scale.
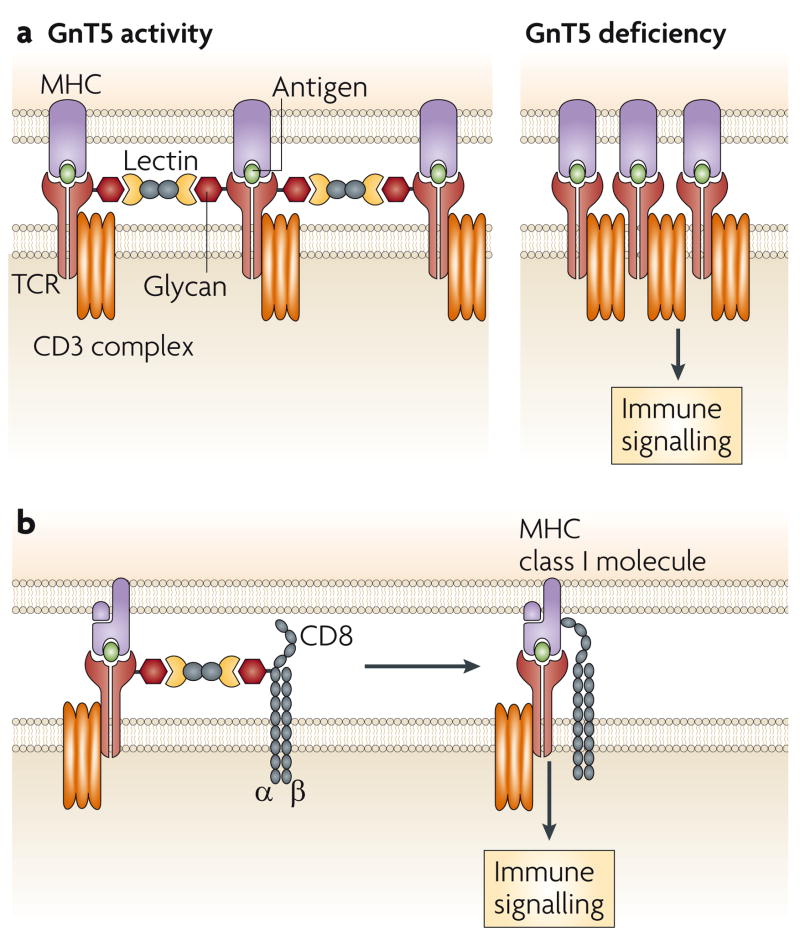
a. Co-clustering of αβ T-cell receptors (TCRs) decreases in the presence of N-glycan branching on the TCR through lectin binding involving one or more of the galectins. Loss of GnT5 glycosyltransferase activity, which mediates N-glycan branching, and therefore lectin binding, increases the propensity for TCR and co-receptor co-clustering and decreases the threshold of TCR activation. b. Lectin binding might also control TCR signalling in cytotoxic T cells by inhibiting the association of the TCR with the CD8 co-receptor.
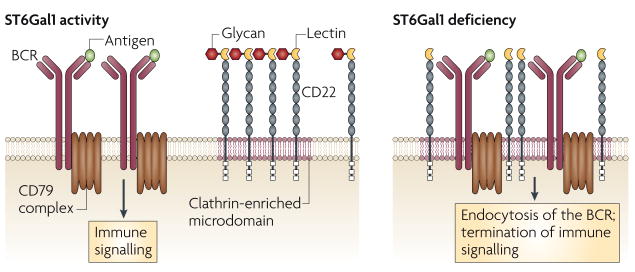
The threshold of B-cell receptor (BCR) activation is modulated by ST6Gal1 and CD22, a member of the Siglec (sialic-acid-binding immunoglobulin-like lectin) family that has a negative role in BCR signalling. CD22 Siglec binding to its glycan ligand promotes homotypic CD22–CD22 interactions, thereby decreasing the frequency of interaction of CD22 with the IgM BCR on mature naive B cells. Loss of CD22 Siglec ligands owing to ST6Gal1 sialyltransferase deficiency markedly increases the co-localization of CD22 with the BCR, thereby increasing the threshold of immune signalling and promoting endocytosis of the BCR in a CD22-dependent manner.
References
-
- Paulson JC, Blixt O, Collins BE. Sweet spots in functional glycomics. Nature Chem Biol. 2006;2:238–248. This publication provides an introduction to the definition and concept of the glycome and describes new approaches to research in this field. - PubMed
-
- Ohtsubo K, Marth JD. Glycosylation in cellular mechanisms of health and disease. Cell. 2006;126:855–867. This article reviews the process of glycosylation and the mechanisms by which glycans contribute to molecular interactions that govern health and disease. - PubMed
-
- Raman R, Raguram S, Venkataraman G, Paulson JC, Sasisekharan R. Glycomics: an integrated systems approach to structure–function relationships of glycans. Nature Methods. 2005;2:817–824. - PubMed
-
- Haltiwanger RS, Lowe JB. Role of glycosylation in development. Annu Rev Biochem. 2004;73:491–537. This article reviews the developmental biology of glycosylation and the mechanisms by which glycans control ontogeny. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
