

Interactions among the transcription factors Runx1, RORgammat and Foxp3 regulate the differentiation of interleukin 17-producing T cells
- PMID: 18849990
- PMCID: PMC4778724
- DOI: 10.1038/ni.1663
Interactions among the transcription factors Runx1, RORgammat and Foxp3 regulate the differentiation of interleukin 17-producing T cells
Erratum in
- Nat Immunol. 2009 Feb;10(2):223
Abstract
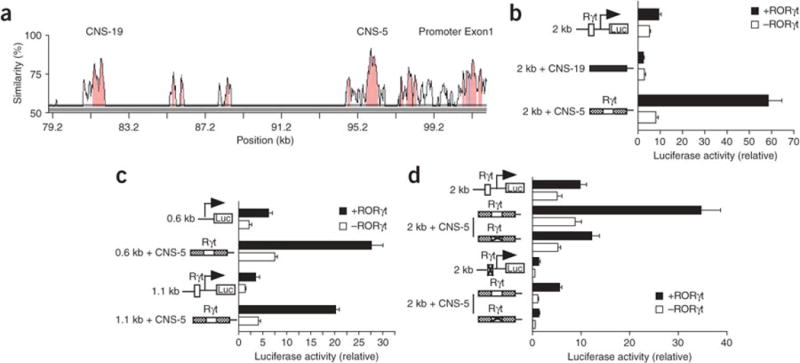
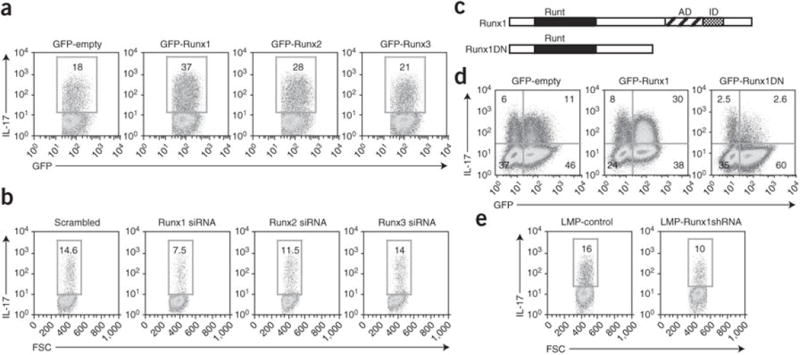
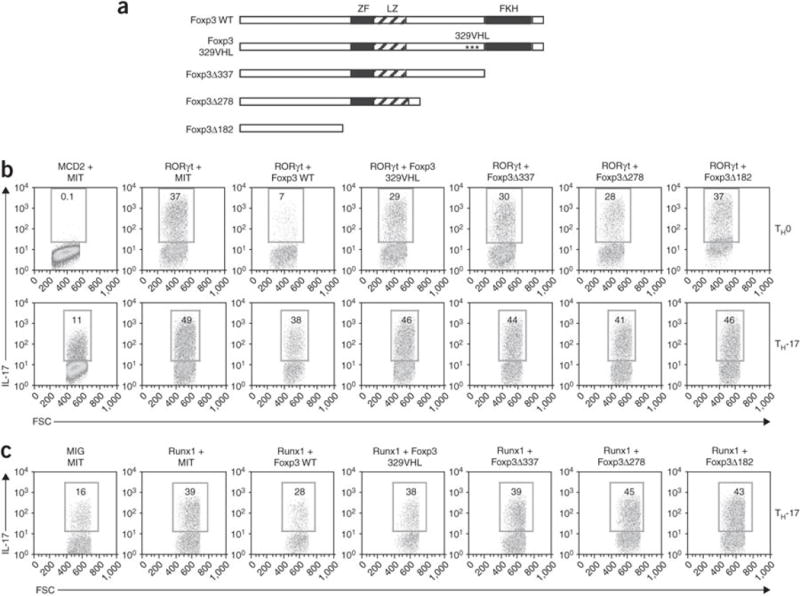
The molecular mechanisms underlying the differentiation of interleukin 17-producing T helper cells (T(H)-17 cells) are still poorly understood. Here we show that optimal transcription of the gene encoding interleukin 17 (Il17) required a 2-kilobase promoter and at least one conserved noncoding (enhancer) sequence, CNS-5. Both cis-regulatory elements contained regions that bound the transcription factors RORgammat and Runx1. Runx1 influenced T(H)-17 differentiation by inducing RORgammat expression and by binding to and acting together with RORgammat during Il17 transcription. However, Runx1 also interacts with the transcription factor Foxp3, and this interaction was necessary for the negative effect of Foxp3 on T(H)-17 differentiation. Thus, our data support a model in which the differential association of Runx1 with Foxp3 and with RORgammat regulates T(H)-17 differentiation.
Figures






References
-
- Ivanov II, et al. The orphan nuclear receptor RORγt directs the differentiation program of proinflammatory IL-17+ T helper cells. Cell. 2006;126:1121–1133. - PubMed
-
- Nakae S, Nambu A, Sudo K, Iwakura Y. Suppression of immune induction of collagen-induced arthritis in IL-17-deficient mice. J Immunol. 2003;171:6173–6177. - PubMed
-
- Komiyama Y, et al. IL-17 plays an important role in the development of experimental autoimmune encephalomyelitis. J Immunol. 2006;177:566–573. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
