

ODA16 aids axonemal outer row dynein assembly through an interaction with the intraflagellar transport machinery
- PMID: 18852297
- PMCID: PMC2568026
- DOI: 10.1083/jcb.200802025
ODA16 aids axonemal outer row dynein assembly through an interaction with the intraflagellar transport machinery
Abstract
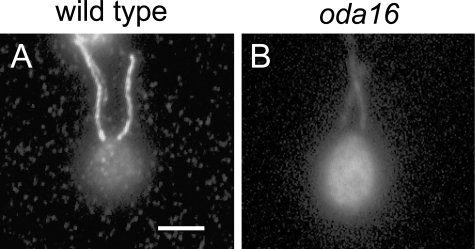
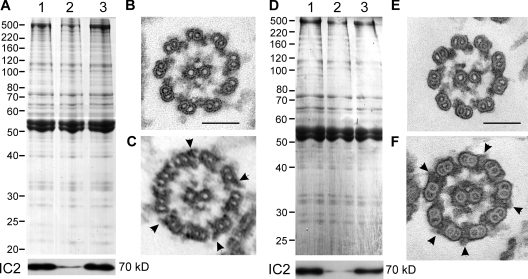
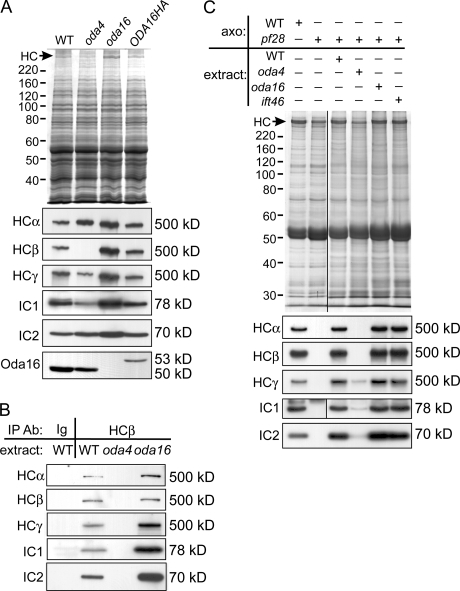
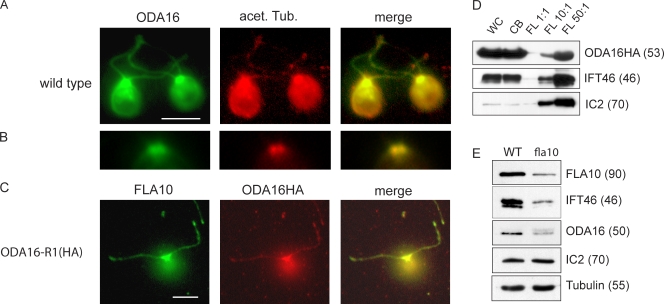
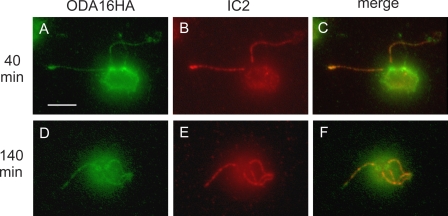
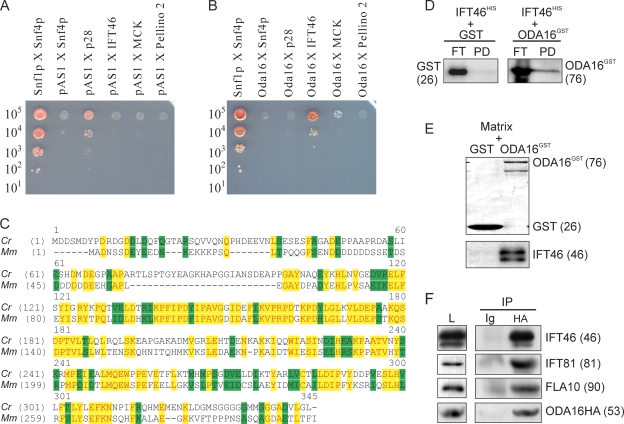
Formation of flagellar outer dynein arms in Chlamydomonas reinhardtii requires the ODA16 protein at a previously uncharacterized assembly step. Here, we show that dynein extracted from wild-type axonemes can rebind to oda16 axonemes in vitro, and dynein in oda16 cytoplasmic extracts can bind to docking sites on pf28 (oda) axonemes, which is consistent with a role for ODA16 in dynein transport, rather than subunit preassembly or binding site formation. ODA16 localization resembles that seen for intraflagellar transport (IFT) proteins, and flagellar abundance of ODA16 depends on IFT. Yeast two-hybrid analysis with mammalian homologues identified an IFT complex B subunit, IFT46, as a directly interacting partner of ODA16. Interaction between Chlamydomonas ODA16 and IFT46 was confirmed through in vitro pull-down assays and coimmunoprecipitation from flagellar extracts. ODA16 appears to function as a cargo-specific adaptor between IFT particles and outer row dynein needed for efficient dynein transport into the flagellar compartment.
Figures
References
-
- Cole, D.G. 2003. The intraflagellar transport machinery of Chlamydomonas reinhardtii. Traffic. 4:435–442. - PubMed
-
- Cole, D.G., D.R. Diener, A.L. Himelblau, P.L. Beech, J.C. Fuster, and J.L. Rosenbaum. 1998. Chlamydomonas kinesin-II-dependent intraflagellar transport (IFT): IFT particles contain proteins required for ciliary assembly in Caenorhabditis elegans sensory neurons. J. Cell Biol. 141:993–1008. - PMC - PubMed
-
- Dutcher, S.K. 2003. Elucidation of basal body and centriole functions in Chlamydomonas reinhardtii. Traffic. 4:443–451. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
