

An inhibitor of gram-negative bacterial virulence protein secretion
- PMID: 18854237
- PMCID: PMC2646588
- DOI: 10.1016/j.chom.2008.08.001
An inhibitor of gram-negative bacterial virulence protein secretion
Abstract
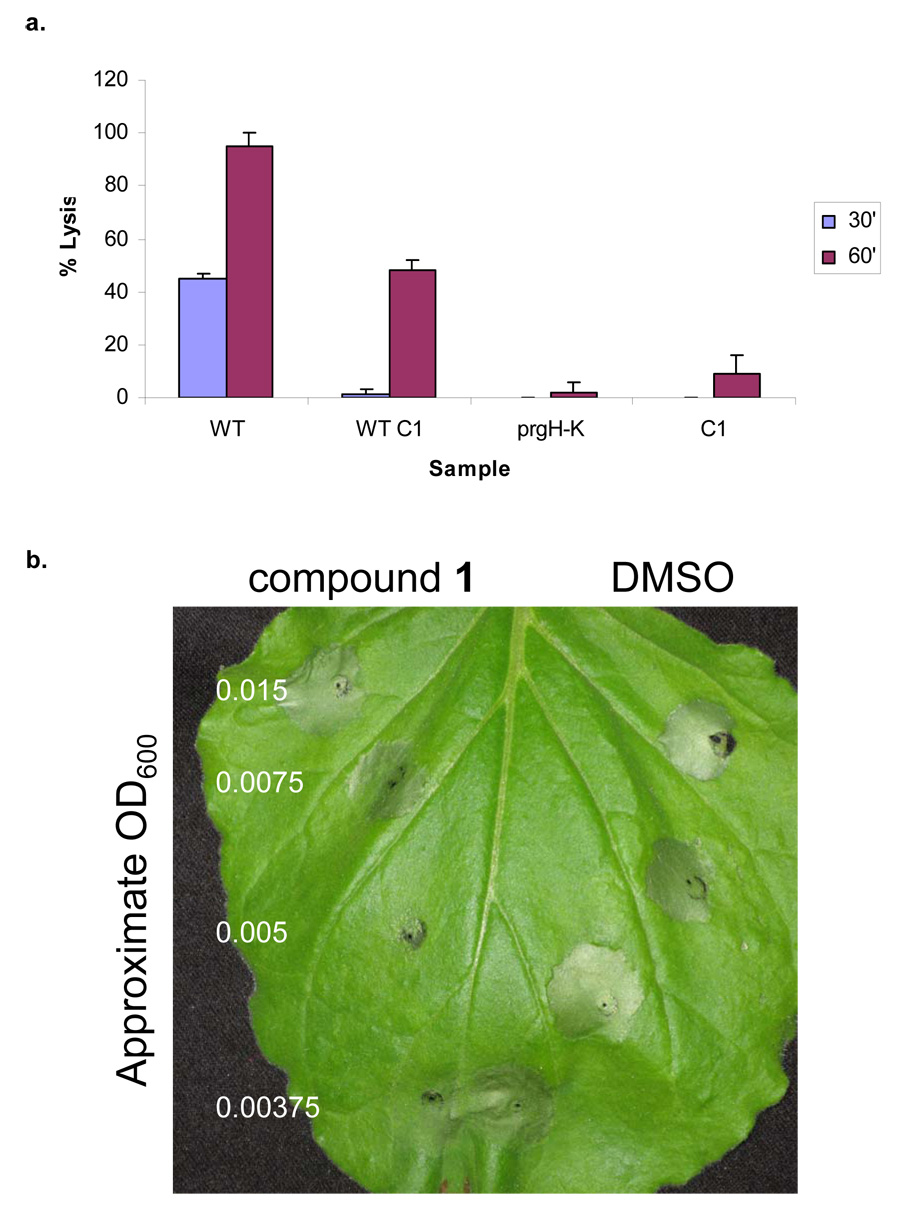
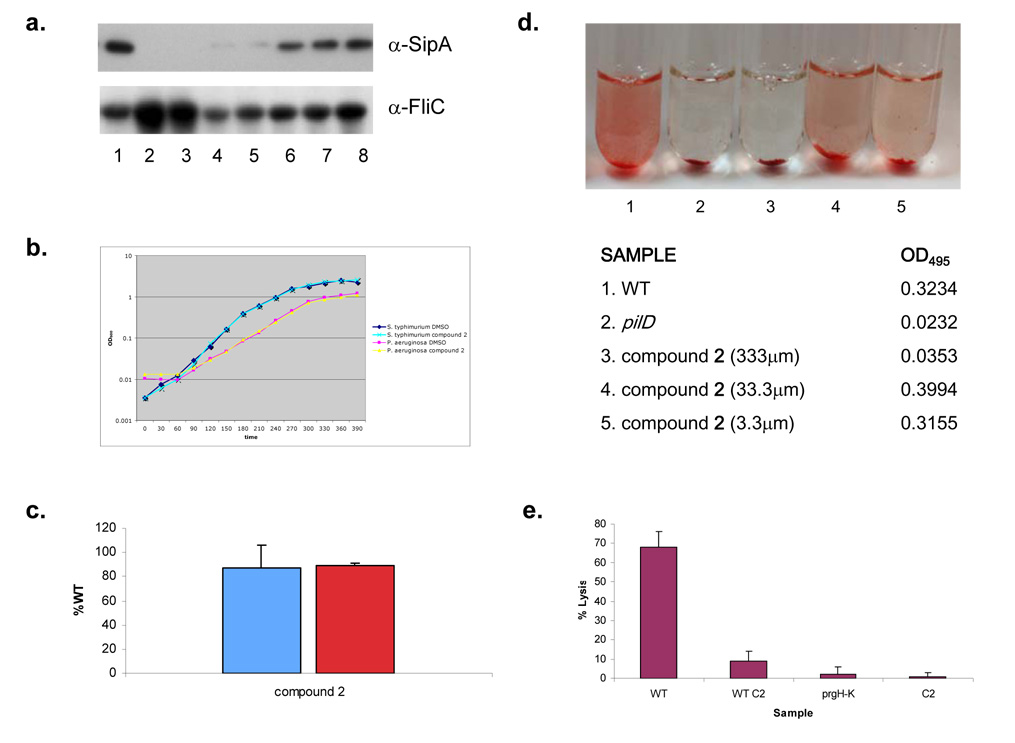
Bacterial virulence mechanisms are attractive targets for antibiotic development because they are required for the pathogenesis of numerous global infectious disease agents. The bacterial secretion systems used to assemble the surface structures that promote adherence and deliver protein virulence effectors to host cells could comprise one such therapeutic target. In this study, we developed and performed a high-throughput screen of small molecule libraries and identified one compound, a 2-imino-5-arylidene thiazolidinone that blocked secretion and virulence functions of a wide array of animal and plant Gram-negative bacterial pathogens. This compound inhibited type III secretion-dependent functions, with the exception of flagellar motility, and type II secretion-dependent functions, suggesting that its target could be an outer membrane component conserved between these two secretion systems. This work provides a proof of concept that compounds with a broad spectrum of activity against Gram-negative bacterial secretion systems could be developed to prevent and treat bacterial diseases.
Figures
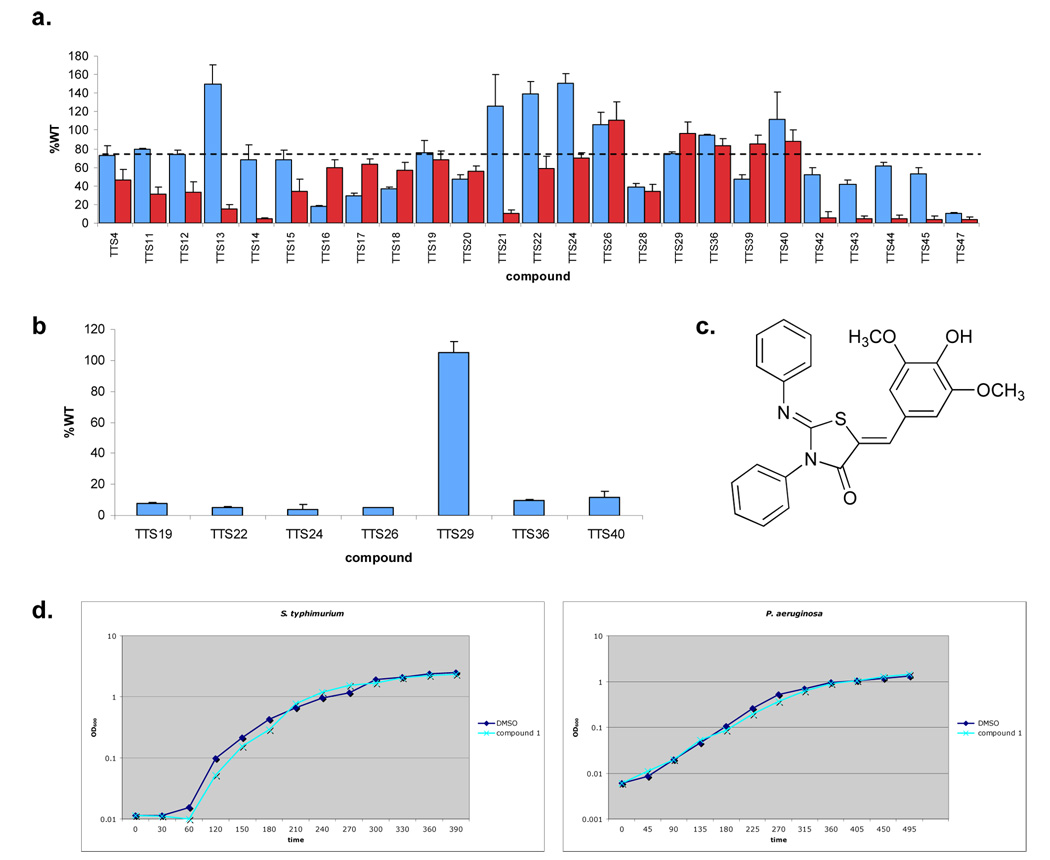
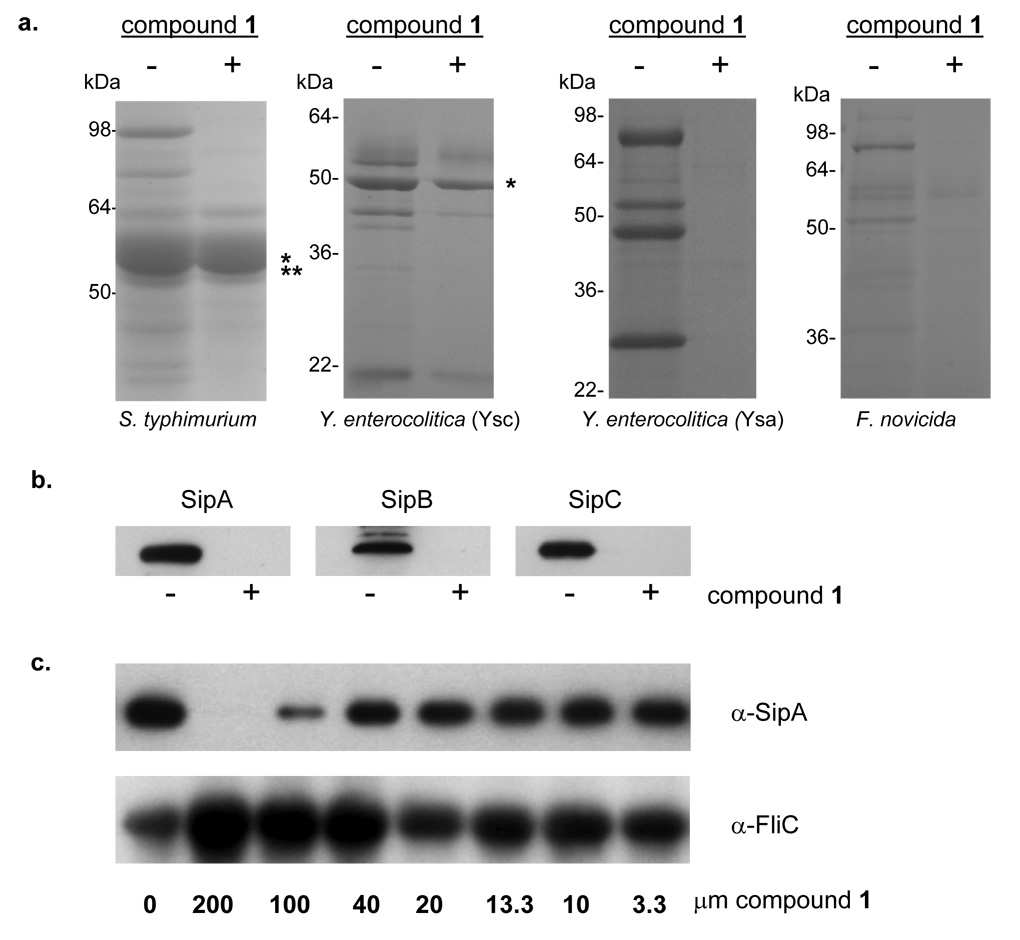
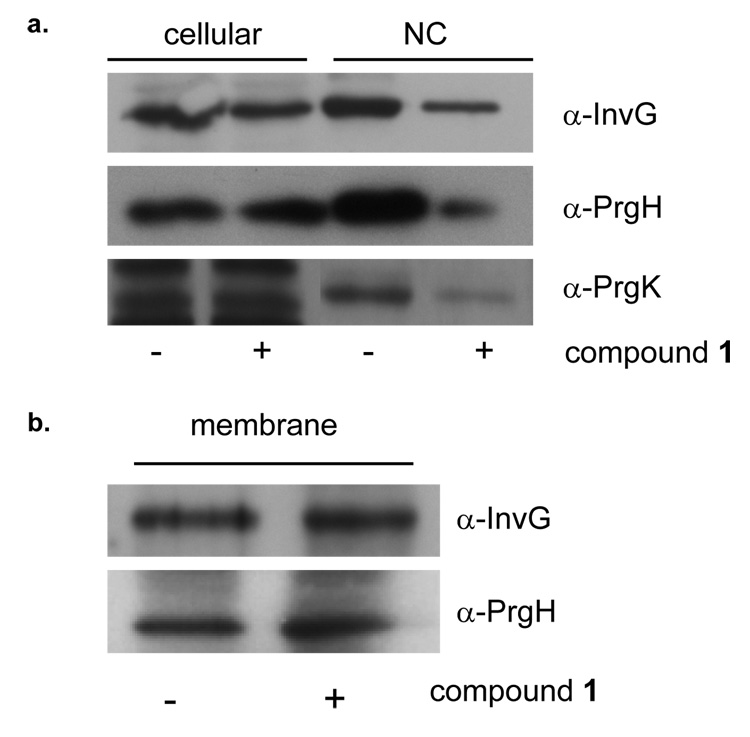
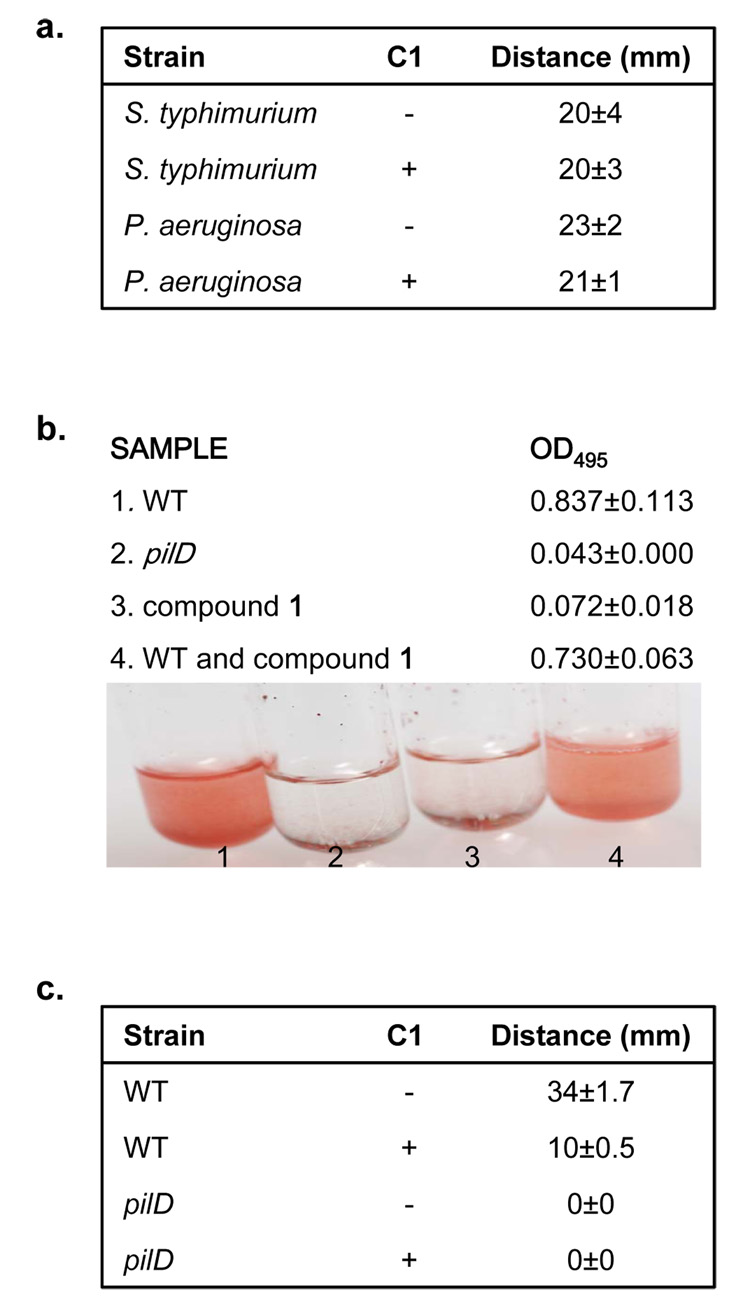
References
-
- Bitter W. Secretins of Pseudomonas aeruginosa: large holes in the outer membrane. Arch Microbiol. 2003;179:307–314. - PubMed
-
- Cianciotto NP. Type II secretion: a protein secretion system for all seasons. Trends Microbiol. 2005;13:581–588. - PubMed
-
- Cornelis GR. The type III secretion injectisome. Nat Rev Microbiol. 2006;4:811–825. - PubMed
-
- Cornelis GR, Van Gijsegem F. Assembly and function of type III secretory systems. Annu Rev Microbiol. 2000;54:735–774. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
