

Molecular phylogeny of a newfound hantavirus in the Japanese shrew mole (Urotrichus talpoides)
- PMID: 18854415
- PMCID: PMC2567236
- DOI: 10.1073/pnas.0808942105
Molecular phylogeny of a newfound hantavirus in the Japanese shrew mole (Urotrichus talpoides)
Abstract
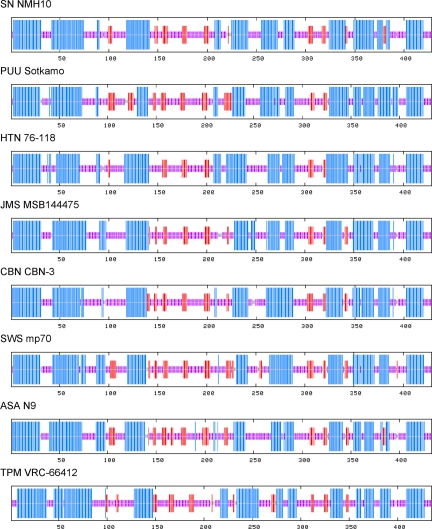
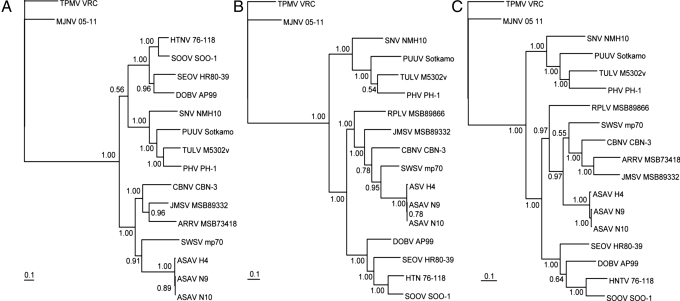
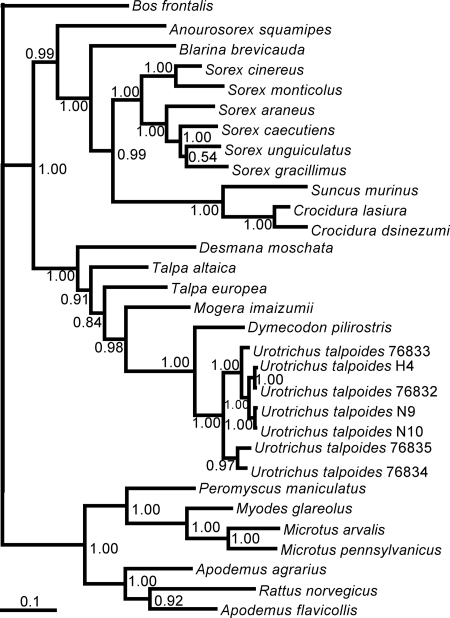
Recent molecular evidence of genetically distinct hantaviruses in shrews, captured in widely separated geographical regions, corroborates decades-old reports of hantavirus antigens in shrew tissues. Apart from challenging the conventional view that rodents are the principal reservoir hosts, the recently identified soricid-borne hantaviruses raise the possibility that other soricomorphs, notably talpids, similarly harbor hantaviruses. In analyzing RNA extracts from lung tissues of the Japanese shrew mole (Urotrichus talpoides), captured in Japan between February and April 2008, a hantavirus genome, designated Asama virus (ASAV), was detected by RT-PCR. Pairwise alignment and comparison of the S-, M-, and L-segment nucleotide and amino acid sequences indicated that ASAV was genetically more similar to hantaviruses harbored by shrews than by rodents. However, the predicted secondary structure of the ASAV nucleocapsid protein was similar to that of rodent- and shrew-borne hantaviruses, exhibiting the same coiled-coil helix at the amino terminus. Phylogenetic analyses, using the maximum-likelihood method and other algorithms, consistently placed ASAV with recently identified soricine shrew-borne hantaviruses, suggesting a possible host-switching event in the distant past. The discovery of a mole-borne hantavirus enlarges our concepts about the complex evolutionary history of hantaviruses.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
-
- Yanagihara R. Hantavirus infection in the United States: Epizootiology and epidemiology. Rev Infect Dis. 1990;12:449–457. - PubMed
-
- Yanagihara R, Gajdusek DC. In: CRC Handbook of Viral and Rickettsial Hemorrhagic Fevers. Gear JHS, editor. Boca Raton: CRC Press; 1988. pp. 151–188.
-
- Lee HW, Lee P-W, Johnson KM. Isolation of the etiologic agent of Korean hemorrhagic fever. J Infect Dis. 1978;137:298–308. - PubMed
-
- Schmaljohn CS, Hasty SE, Harrison SA, Dalrymple JM. Characterization of Hantaan virions, the prototype virus of hemorrhagic fever with renal syndrome. J Infect Dis. 1983;148:1005–1012. - PubMed
-
- Brummer-Korvenkontio M, et al. Nephropathia epidemica: Detection of antigen in bank voles and serologic diagnosis of human infection. J Infect Dis. 1980;141:131–134. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
