

Review
doi: 10.1038/nrn2501.
Epub 2008 Oct 15.
Silent synapses and the emergence of a postsynaptic mechanism for LTP
Affiliations
- PMID: 18854855
- PMCID: PMC2819160
- DOI: 10.1038/nrn2501
Item in Clipboard
Review
Silent synapses and the emergence of a postsynaptic mechanism for LTP
Nat Rev Neurosci.
2008 Nov.
Erratum in
- Nat Rev Neurosci. 2009 Mar;10(3):242
Abstract
Silent synapses abound in the young brain, representing an early step in the pathway of experience-dependent synaptic development. Discovered amidst the debate over whether long-term potentiation reflects a presynaptic or a postsynaptic modification, silent synapses--which in the hippocampal CA1 subfield are characterized by the presence of NMDA receptors but not AMPA receptors--have stirred some mechanistic controversy of their own. Out of this literature has emerged a model for synapse unsilencing that highlights the central role for postsynaptic AMPA-receptor trafficking in the expression of excitatory synaptic plasticity.
Figures
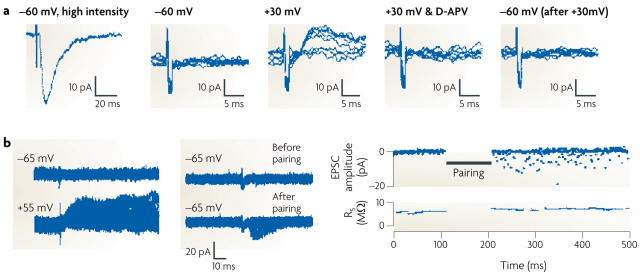
A silent synapse is defined as a synapse in which an excitatory postsynaptic current (EPSC) is absent at the resting membrane potential but becomes apparent on depolarization. The traces here were obtained during whole-cell recordings from CA1 pyramidal neurons from acute rat hippocampal slices. a | (From left to right.) High-intensity stimulation evoked a fast, AMPAR-mediated EPSC at a holding potential of −60 mV. When the stimulus intensity was reduced to below the threshold for triggering EPSCs, as shown in a series of superimposed traces from repeated trials, no evoked current appeared at −60 mV. However, using the same stimulus intensity, a slow EPSC did appear at a holding potential of +30 mV; this EPSC disappeared on application of the NMDAR antagonist D-APV, indicating that it was purely mediated by NMDARs. On returning the holding potential to −60 mV, the lower-intensity stimulus again did not evoke a current. The flat traces in this series indicate failures (see BOX 2). b | The left-hand panel shows an EPSC appearing at baseline at a holding potential of +55 mV but not at −65 mV. However, after a long-term potentiation protocol, in which stimulation was paired with postsynaptic depolarization to 0 mV, EPSCs appeared at −65 mV (middle panel). As illustrated in the time course graph (right-hand panel), the number of failures diminished markedly after pairing. Part a reproduced, with permission, from REF. © (1995) Elsevier Science. Part b reproduced, with permission, from REF. © (1995) Macmillan Publishers Ltd. All rights reserved.
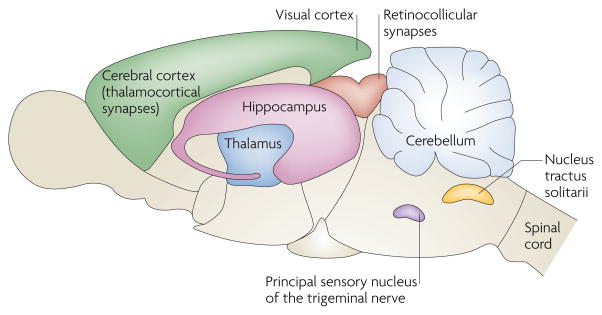
Since the original discovery of silent synapses in the hippocampus,,, such synapses have now been described all over the CNS. In rats, silent synapses are abundant at thalamocortical connections during postnatal days (P) 2–5 and are markedly reduced by P8–9 (REF. 122). Silent synapses exhibit a similar developmental gradient in the rat visual cortex, being easily detectable at P3–5 and significantly reduced by P9–11 (REFS 123,124). At retinocollicular connections in mice, and at cerebellar granule cells, nucleus tractus solitarii neurons and the dorsal horn of the spinal cord, in rats, the prevalence of silent synapses declines over the first two postnatal weeks. Interestingly, in rats, synapses in the principal sensory nucleus of the trigeminal nerve have matured by birth, but they can be silenced by postnatal deafferentation. At all of these anatomical locations, as in the hippocampal CA1 subfield, silent synapses were signified by the presence of an NMDAR-mediated excitatory postsynaptic current (EPSC) and the absence of AMPAR-mediated EPSCs. Silent synapses and their unsilencing seem to be pervasive features of excitatory transmission, contributing to synaptic development and plasticity.
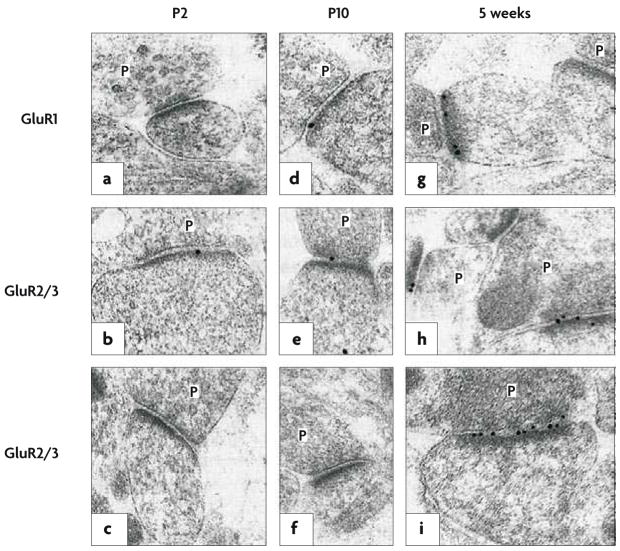
In hippocampal tissue obtained from young rats, postembedding immunogold labelling was performed using an antibody raised against the carboxyl terminus of the AMPAR subunit GluR1 (a,d,g) or using antibodies that bind to both the GluR2 and the GluR3 subunits (b,c,e,f,h,i); both GluR1 and GluR2 contribute to virtually every tetrameric AMPAR complex in CA1 pyramidal cells. The age at which the tissue was obtained increases from left to right, from postnatal day 2 (P2) to 5 weeks. In each part of the figure, the presynaptic terminals, with glutamate-containing vesicles, are labelled P; opposite each of these terminals is the postsynaptic membrane, marked by the presence of an electron-dense band (the postsynaptic density) where glutamate receptors (black dots) crowd alongside associated anchoring and signalling proteins. A large increase in AMPAR-subunit labelling, relative to the postnatal period, is evident at 5 weeks. Figure reproduced, with permission, from REF. © (1999) Macmillan Publishers Ltd. All rights reserved.
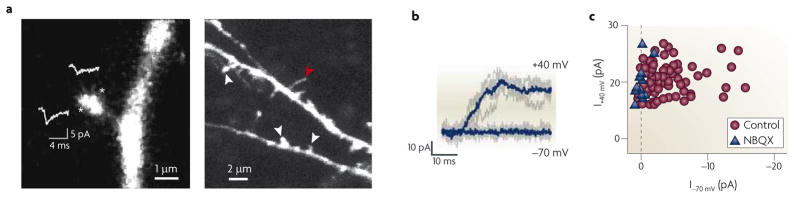
a | The left-hand panel shows a spine and its parent dendrite, with sites of glutamate uncaging indicated by asterisks; the show the average| AMPAR-mediated unitary excitatory postsynaptic current (uEPSC) at each site. On each spine tested, an area of maximal response (in this case the site indicated by the lower asterisk) could be located, illustrating the precision of the technique of glutamate uncaging. The right-hand panel shows a lower-magnification view of the basal dendrite of a layer II/III cortical pyramidal neuron from a postnatal day (P) 9 rat, showing examples of mature spines (indicated by the white arrowheads) and a filopodium (indicated by the red arrowhead). b | At P9, glutamate uncaging at a spine revealed no uEPSC at −70 mV but did reveal a slow, NMDAR-mediated uEPSC at +40 mV; five individual trials (grey traces) and the corresponding averages (blue traces) are shown. This is a silent synapse that clearly lacks functional AMPARs in the postsynaptic membrane. c | NMDAR-mediated uEPSC amplitudes versus AMPAR-mediated uEPSC amplitudes are plotted in control conditions (n = 72 spines, P9–13) or in the presence of the AMPAR antagonist NBQX (n = 13 spines, P9–15). The dashed line marks the expected position for silent spines lacking AMPARs. Many control spines fall along this line, exhibiting AMPAR-mediated uEPSCs similar to those recorded in the presence of NBQX, illustrating the large number of silent synapses in these young animals. During the second postnatal week, silent synapses became increasingly hard to find and had virtually disappeared by P14–17 (not shown). Parts a and b reproduced and modified (respectively), with permission, from REF. © (2008) Cambridge University Press.
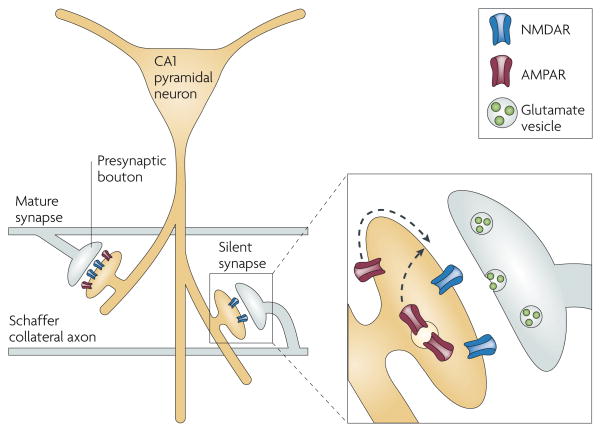
A CA1 pyramidal neuron projects its large apical dendrite down into the stratum radiatum, where Schaffer collateral axons en passant synapses onto dendritic spines. The spine pictured on the left is mature, with a full complement of both AMPARs and NMDARs. By contrast, the spine on the right possesses only NMDARs and therefore cannot conduct current in response to presynaptic glutamate release. The expanded view of this silent synapse illustrates how an LTP-induction protocol will cause AMPARs to migrate towards the postsynaptic density, either through lateral diffusion along the synaptic membrane or through the fusion of AMPAR-containing endosomes,,.
References
-
- Malenka RC, Kauer JA, perkel DJ, Nicoll RA. The impact of postsynaptic calcium on synaptic transmission — its role in long-term potentiation. Trends Neurosci. 1989;12:444–450. - PubMed
-
- Liao D, Hessler NA, Malinow R. Activation of postsynaptically silent synapses during pairing-induced LTP in CA1 region of hippocampal slice. Nature. 1995;375:400–404. This paper and reference 3, which was published two months later, first established the presence of hippocampal CA1 silent synapses. Both groups used a minimal-stimulation technique to isolate synapses exhibiting the (now classic) physiological signature of a silent synapse — the presence of N-EPSCs but not A-EPSCs — and both groups went on to show that LTP triggered the rapid recruitment of an AMPAR-mediated response. - PubMed
-
- Isaac JT, Nicoll RA, Malenka RC. Evidence for silent synapses: implications for the expression of LTP. Neuron. 1995;15:427–434. - PubMed
-
- Wall PD. The presence of ineffective synapses and the circumstances which unmask them. Philos Trans R Soc Lond B Biol Sci. 1977;278:361–372. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
