

Noradrenergic induction of odor-specific neural habituation and olfactory memories
- PMID: 18923046
- PMCID: PMC2588668
- DOI: 10.1523/JNEUROSCI.3853-08.2008
Noradrenergic induction of odor-specific neural habituation and olfactory memories
Abstract
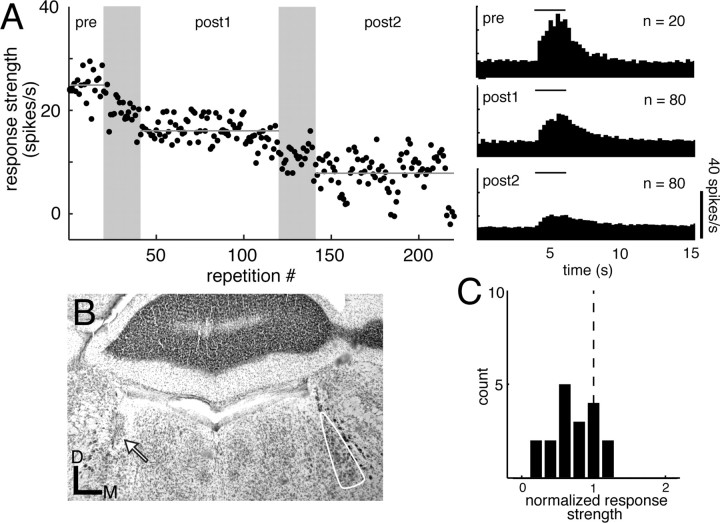
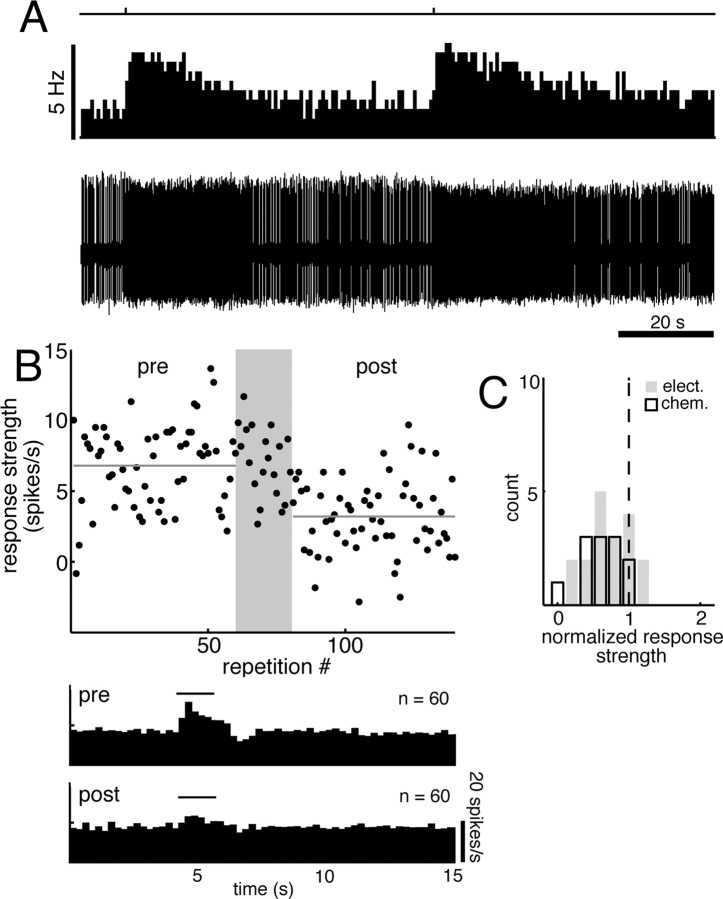
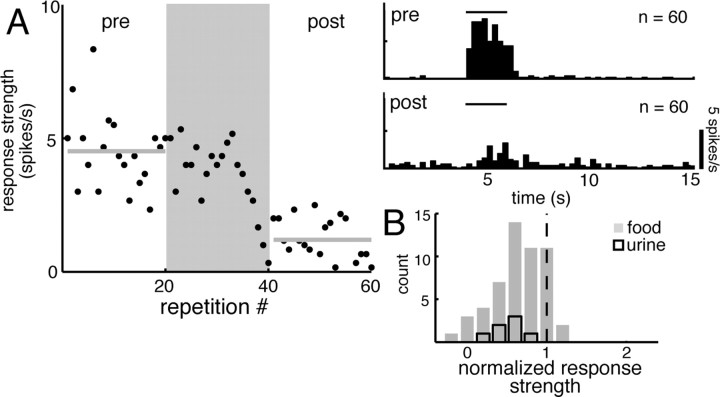
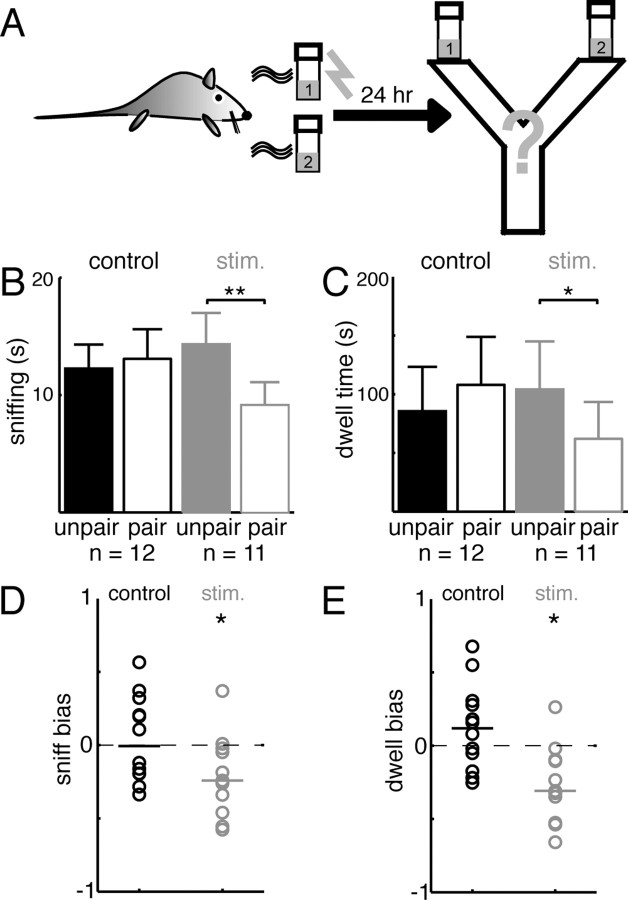
For many mammals, individual recognition of conspecifics relies on olfactory cues. Certain individual recognition memories are thought to be stored when conspecific odor cues coincide with surges of noradrenaline (NA) triggered by intensely arousing social events. Such familiar stimuli elicit reduced behavioral responses, a change likely related to NA-dependent plasticity in the olfactory bulb (OB). In addition to its role in these ethological memories, NA signaling in the OB appears to be relevant for the discrimination of more arbitrary odorants as well. Nonetheless, no NA-gated mechanism of long-term plasticity in the OB has ever been directly observed in vivo. Here, we report that NA release from locus ceruleus (LC), when coupled to odor presentation, acts locally in the main OB to cause a specific long-lasting suppression of responses to paired odors. These effects were observed for both food odors and urine, an important social recognition cue. Moreover, in subsequent behavioral tests, mice exhibited habituation to paired urine stimuli, suggesting that this LC-mediated olfactory neural plasticity, induced under anesthesia, can store an individual recognition memory that is observable after recovery.
Figures


References
-
- Adams LM, Foote SL. Effects of locally infused pharmacological agents on spontaneous and sensory-evoked activity of locus coeruleus neurons. Brain Res Bull. 1988;21:395–400. - PubMed
-
- Aston-Jones G, Cohen JD. An integrative theory of locus coeruleus-norepinephrine function: adaptive gain and optimal performance. Annu Rev Neurosci. 2005;28:403–450. - PubMed
-
- Berridge CW, Abercrombie ED. Relationship between locus coeruleus discharge rates and rates of norepinephrine release within neocortex as assessed by in vivo microdialysis. Neuroscience. 1999;93:1263–1270. - PubMed
-
- Berridge CW, Waterhouse BD. The locus coeruleus-noradrenergic system: modulation of behavioral state and state-dependent cognitive processes. Brain Res Brain Res Rev. 2003;42:33–84. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous