

Analysis of prelamin A biogenesis reveals the nucleus to be a CaaX processing compartment
- PMID: 18923140
- PMCID: PMC2592638
- DOI: 10.1091/mbc.e08-07-0704
Analysis of prelamin A biogenesis reveals the nucleus to be a CaaX processing compartment
Abstract
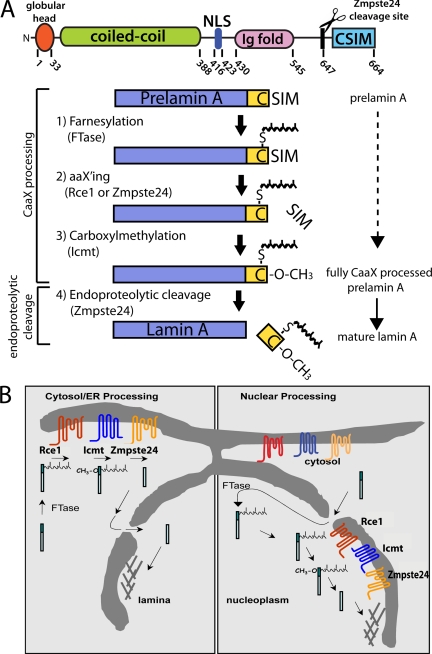
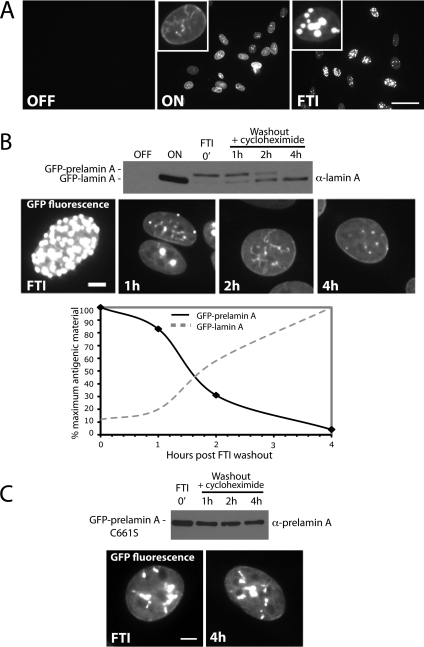
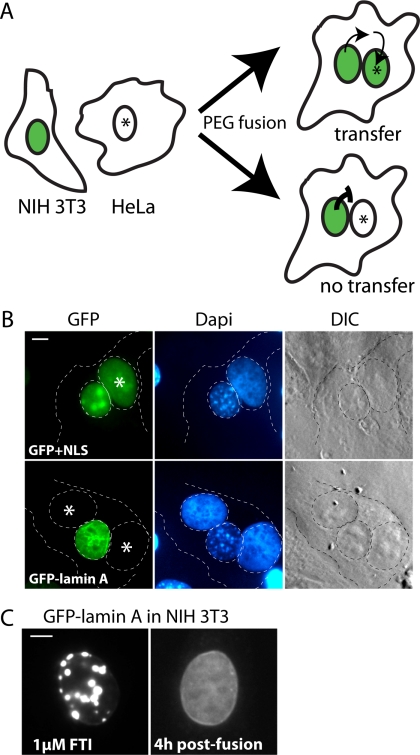
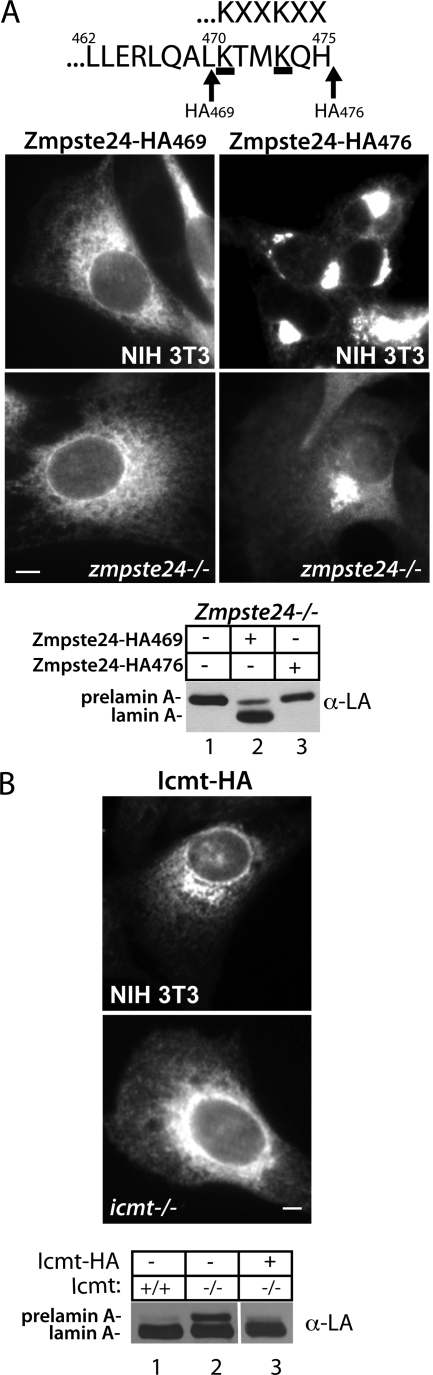
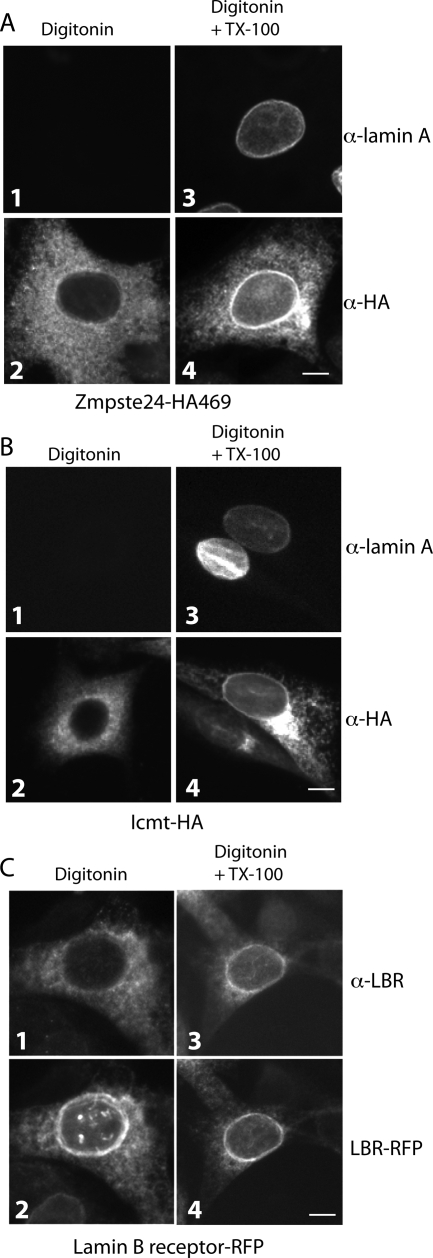
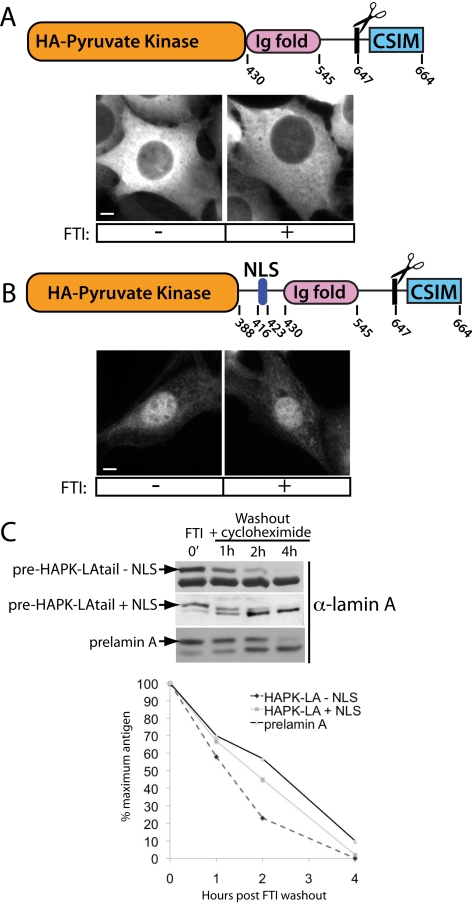
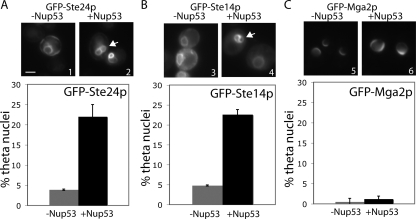
Proteins establish and maintain a distinct intracellular localization by means of targeting, retention, and retrieval signals, ensuring most proteins reside predominantly in one cellular location. The enzymes involved in the maturation of lamin A present a challenge to this paradigm. Lamin A is first synthesized as a 74-kDa precursor, prelamin A, with a C-terminal CaaX motif and undergoes a series of posttranslational modifications including CaaX processing (farnesylation, aaX cleavage and carboxylmethylation), followed by endoproteolytic cleavage by Zmpste24. Failure to cleave prelamin A results in progeria and related premature aging disorders. Evidence suggests prelamin A is imported directly into the nucleus where it is processed. Paradoxically, the processing enzymes have been shown to reside in the cytosol (farnesyltransferase), or are ER membrane proteins (Zmpste24, Rce1, and Icmt) with their active sites facing the cytosol. Here we have reexamined the cellular site of prelamin A processing, and show that the mammalian and yeast processing enzymes Zmpste24 and Icmt exhibit a dual localization to the inner nuclear membrane, as well as the ER membrane. Our findings reveal the nucleus to be a physiologically relevant location for CaaX processing, and provide insight into the biology of a protein at the center of devastating progeroid diseases.
Figures
References
-
- Ashar H. R., James L., Gray K., Carr D., Black S., Armstrong L., Bishop W. R., Kirschmeier P. Farnesyl transferase inhibitors block the farnesylation of CENP-E and CENP-F and alter the association of CENP-E with the microtubules. J. Biol. Chem. 2000;275:30451–30457. - PubMed
-
- Capell B. C., Collins F. S. Human laminopathies: nuclei gone genetically awry. Nat. Rev. Genet. 2006;7:940–952. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
